Download

1 / 36
480 likes | 1.14k Views
Metabolism of Red Blood Cell & Membrane Stability. Prof Dr Kwan Teck Kim Department of Biochemistry Faculty of Medicine Mahsa University College. Learning outcomes. On completion of this topic, the student will be able to:

E N D
Metabolism of Red Blood Cell&Membrane Stability Prof Dr Kwan Teck Kim Department of Biochemistry Faculty of Medicine Mahsa University College
Learning outcomes On completion of this topic, the student will be able to: • Discuss the role of glycolysis in erythrocytes with respect to ATP production, supply of intermediates for Rapoport-Lueberingglycolytic shunt and Hexosemonophosphate pathway. • Explain the role of NADPH in glutathione metabolism • Explain the role of ATP and glutathione in the maintenance of erythrocyte membrane stability • Explain the molecular basis of hemolytic anemias
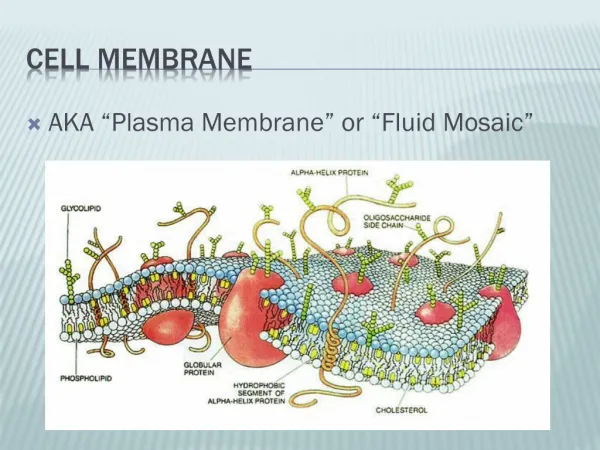
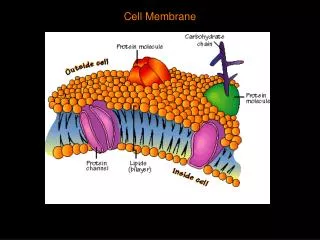
Red Blood Cell • The red blood cell is simple in terms of its structure and function, consisting principally of a concentrated solution of hemoglobin surrounded by a membrane. The erythrocyte membrane is flexible because of its cytoskeletal structure. • The red cell contains a battery of cytosolic enzymes, such as suproxide dismutase, catalase, and glutathione peroxidase, to dispose of powerful oxidants (ROS, reactive oxygen species) generated during its metabolism. • Genetically determined deficiency of the activity of glucose-6-phosphate dehydrogenase, which produces NADPH, is an important cause of hemolytic anemia.
Rapoport-Lueberingglycolytic shunt • To generate 2,3-Bisphosphoglycerate (2,3-BPG) • 2,3-BPG is important regulator of binding of O2 to Hb. • HbO2+ 2,3-BPG ↔ Hb-2,3-BPG + O2 (deoxyhemoglobin)
2,3-Bisphosphoglycerate shunt ADP ATP 2-
2,3-Bisphosphoglycerate mutase/phosphatase • This enzyme is bifunctional, serving as mutase for the formation of 2,3-bisphosphoglycerate and also as a phosphatase. • Phosphatase hydrolyzes 2,3-bisphosphoglycerate to 3-phosphoglycerate and Pi . • Erythrocytes contain very high concentrations of 2,3-bisphosphoglycerate, which functions as important negative allosteric regulator of the binding of oxygen to hemoglobin.
2,3-Bisphosphoglycerate mutase/phosphatase (contd) • 15-25% of glucose converted to lactate in red blood cells goes by way of the “BPG shunt”for the synthesis of 2,3-bisphosphoglycerate. • No net production of ATP occurs when glucose is converted to lactate by the BPG shunt as it bypasses the phosphoglyceratekinase step. • However, it does serve to provide 2,3-BPG, which binds to Hb, decreasing its affinity for O2, and so making O2 more readily available to tissues.
2,3-Bisphosphoglycerate (BPG) in Red Blood Cells Modulates Oxygen release from Hemoglobin • BPG dissociates as deoxy-Hb is converted to oxy-Hb. H+BPG + Hb + 4O2 ↔ Hb(O2)4 + BPG + nH+ • Increased concns of BPG force the equilibrium to the left, and they correspondingly shift the saturation plot to the right with an increase in P50 (see Fig. 9.34). • In contrast, lowered concns of BPG force this equilibrium to the right, & correspondingly shift the saturation plot to the left with a lower P50 .
2,3-Bisphosphoglycerate (BPG) in Red Blood Cells Modulates Oxygen release from Hemoglobin (contd 1) • BPG is formed in a minor pathway for glucose metabolism & is present in small amounts in all cells. • In rbc, this pathway is highly active, & BPG concns are approx. equimolar to that of Hb. • BPG binds to the deoxy (T) but not to oxy (R) conformation. • Binding of BPG to Hb stabilizes the T conformation & increases its concn relative to the R conformation.
2,3-Bisphosphoglycerate (BPG) in Red Blood Cells Modulates Oxygen release from Hemoglobin (contd 2) • Conditions that cause hypoxia (deficiency of oxygen) such as anemia, smoking, and high altitude increase BPG levels in rbc. • In turn, conditions leading to hyperoxia result in lower levels of BPG. • Changes in rbc levels of BPG are slow & occur over hours and days to compensate for chronic changes in pO2 levels.
Glucose transport & metabolism • Depending on the concn gradient, erythrocyte GLUT 1 (glucose transporter) facilitates transport of glucose in both directions • Glucose is metabolized mainly by glycolysis in erythrocytes. • Since erythrocytes lack mitochondria, the end product of glycolysis is lactic acid, which is released into the blood plasma.
Glucose metabolism via PPP • Glucose used by the Pentose Phosphate pathway in r.b.c. provides NADPH to keep glutathione in the reduced state, which has an important role in the destruction of organic peroxides and H2O2. • Peroxides cause irreversible damage to membranes, DNA and other cellular components and must be removed to prevent cell damage and death.
Erythrocyte and Pentose Phosphate Pathway • Glucose 6-phosphate dehyrogenase deficiency causes drug induced hemolyticanemia • Patients administered the drug pamaquine experienced drastic loss of Hb • Role of NADPH in RBC’s is to reduce gluthathione to sulfhydryl form • Essential for maintaining structure of Hb (keeps Hb in ferrous state) • [ G-S-S-G → 2G-SH ]
Energy generation • Generates ATP for energy to maintain • RBC shape and flexibility • Via anaerobic glycolytic pathway • 2 ATP for each molecule glucose
Overview of erythrocyte metabolism • Mature erythrocytes contain no intracellular organelles, so metabolic enzymes of rbc are limited to those found in cytoplasm. • Glycolysisis the major pathway, with branches for the hexosemonophosphate shunt (for protection against oxidizing agents) and • Rapoport-Luebering shunt (which generates 2,3-BPG, which moderates oxygen binding to Hb). • The NADH generated from glycolysis can be used to reduce methemoglobin (Fe3+) to normal Hb (Fe2+) or • To convert pyruvate to lactate, so that NAD+ can be regenerated and used for glycolysis.
RBC Membrane Integrity • Glutathione is present in RBCs. This is used for inactivation of free radicals formed inside RBC. • The enzyme used is glutathione peroxidase. • The glutathione is regenerated by an NADPH dependent glutathione reductase. • The NADPH is derived from the glucose 6-phosphate dehydrogenase shunt pathway. • The occurrence of hemolysis in G6PD deficiency is attributed to the decreased regeneration of reduced glutathione.
Pentose phosphate pathway (Hexosemonophosphate shunt) 6-Phosphogluco- lactonase C5 + C5 ↔ C3 + C7 Transketolase C7 + C3 ↔ C4 + C6 Transaldolase C5 + C4 ↔ C3 + C6 Transketolase 6-Phosphogluconate dehydrogenase Oxidative phase ← Non-Oxidative Phase ←
Transketolase & transaldolase reactions • C5 + C5 <---------------> C3 + C7 (transketolase) • C7 + C3 <---------------> C4 + C6 (transaldolase) • C5 + C4 <----------------> C3 + C6 (transketolase) • Transketolase- transfers 2 carbon units. • Transaldolase – transfers 3 carbon units.
Pentose Phosphate Pathway • The oxidative branch consists of 3 irreversible reacns. • Glucose-6-phosphate dehydrogenase (G6PD), the rate-limiting enzyme, converts glucose 6-phosphate to 6-phosphogluconolactone, which is further converted in a series of reacns to ribulose 5-phosphate. • The 2 NADPH formed are used for reductive biosynthesis and for maintaining the anti-oxidant glutathione in the reduced form.
Pentose Phosphate Pathway (contd) • All of the reacns of the non-oxidative branch are reversible and produce ribose 5-phosphate for RNA and DNA synthesis. • Also produced are the intermediates fructose 6-phosphate and glyceraldehde 3-phosphate for glycolysis (fed state) or gluconeogenesis (fasting state).
Effect of H+ on oxygen binding by Hb • A. In the tissues, CO2 is released. In rbc, this CO2 forms carbonic acid, which releases protons. The protons bind to Hb, causing it to release oxygen to the tissues. • B. In the lungs, the reacns are reversed. O2 binds to protonatedHb, causing the release of protons. They bind to HCO3- , forming carbonic acid, which is cleaved to water and CO2 , which is exhaled.
References • Lieberman, M. & Marks, A. (2009). Marks’ Basic Medical Biochemistry: A clinical approach (3rd edition), Lippincott, Williams & Wilkins, Philadelphia. • Harvey, R.A. & Ferrier, D.R. (2011). Lippincott’s Illustrated Reviews: Biochemistry (5th edition), Lippincott, Williams and Wilkins, Philadelphia. • Vasudeven, D.M., Sreekumari, S. & Vaidyanathan, K. (2011). Textbook of Biochemistry for Medical Students (6th edition), JaypeeBrothrs Medical Publishers Ltd, New Delhi.