Download

1 / 15
150 likes | 256 Views
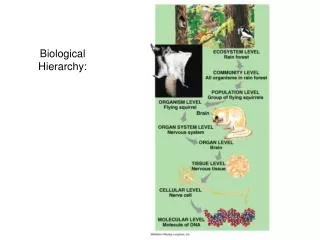
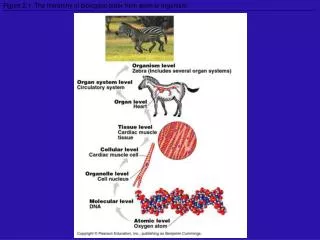
Two notions of biological hierarchy. John Collier University of KwaZulu-Natal collierj@ukzn.ac.za. Typical Biological Hierarchy. Physical Chemical Macromolecules (especially genetic) Cells Organs Organisms Populations Ecologies Species Higher Taxa.

E N D
Two notions of biological hierarchy John Collier University of KwaZulu-Natal collierj@ukzn.ac.za
Typical Biological Hierarchy Physical Chemical Macromolecules (especially genetic) Cells Organs Organisms Populations Ecologies Species Higher Taxa ISHPSSB Salt Lake City 2011
Two types of hierarchies • Class, types • Real or nominal • Hierarchy type fits all levels of hierarchy and determines containment, e.g., substance, function, science • Component, spatial containment, token • Real of nominal (e.g., things vs. aggregates – Wimsatt, dynamical systems vs. sets) • Whole – component relation determines containment • These two kinds of hierarchies are sometimes unconsciously conflated (e.g. the physical components of something are referred to as its physical level, and then sometimes this slides into discussion of type properties of the physical, applied back to discussion of the components in terms of a type level)
Two types of hierarchies - Classes • Types, classes • Called determinable-determinate by W.E. Johnson (1921) • Called specification hierarchy by Stan Salthe (1991) • Salthe’s formalism: {physical {chemical {cells {organs {organisms}}}}} • Another version: {physical {thermodynamic {self-organized {living {mental {social}}}}
Two types of hierarchies - Individuals • Individuals, things, tokens, whole-part, spatial (Salthe) • I prefer not to use “token” as token as generally thought of being defined under classes, but real individuals have their own defining characteristics, and exist independently of any classes or types (more soon) • Types are an abstraction from individuals (Collier) • Real individuals defining characteristic is their cohesion (Collier) • I will concentrate on these as they are less understood by philosophers than class hierarchies.
Cohesion • Unity relation (Perry): What makes parts of a thing parts of the same thing, closure of part relation. Depends on type of thing. • For real objects and properties the unity relation is always dynamical (Dynamical realism, Hooker, Collier) • Essential to the existence of the thing, determines its Lockeanreal essence if it is an individual (e.g., Lockean account of personal identity in terms of memory transfer) • Cohesion comes in degrees, so it is not always clear cut where boundaries lie.
Cohesion in individual hierarchies • Each level has cohesion of its own; it is not merely an aggregate (Collier Entropy, 2003) • E.g., cells have their own cohesion • This constrains the activities of their chemical components (not to mention, of their immediate environment – cohesive closure is not complete, and may extend beyond the usual boundaries of the cohesive entity) • Cohesion exerts downward causation • This is not simply the effect of the lower level components (Paul Humphreys: fusion)
Cohesion in biology 1 • In biology cohesion organizational, and self-maintaining (Maturana – autopoiesis, Rosen – closure to efficient causation) • But contrary to Maturana and Rosen, and consistent with General Systems Theory (Bertalanffy) biological cohesion is open in all respects (including information and effcient causation). This is a consequence of the fact that cohesion comes in degrees. • Cohesive closure can involve environmental loops and is open to new environmental inputs that are not system predictable (Collier, CASYS 2003). The latter can cause reorganization of the system around these properties (systemic adaptation).
Cohesion in biology 2 • Cohesive levels form from lower levels, sometimes through self-organization, which results in emergence (non-reducibility) • Sometimes levels form between previous levels, constrained dynamical from both above and below. • Interaction of parts at a given type-level may or may not be strong enough to be cohesive enough to produce a new level. • Hypothesis – new individual levels in biological systems are primarily organization, and have autonomy, which is the organizational property that produces self-persistence. • Autonomy is the primary form of cohesion in biological systems.
Obligatory connection to species • Cohesion was introduced by E.O. Wiley in PhylogeneticSystematics in the form of horizontal and vertical cohesion of species. • I extended this notion to all dynamical entities in part to justify speculative work of Brooks and Wiley on dynamical biological information systems. • Species are cohesive. Ecological factors are one of several factors in species cohesion. • As will follow from what I say next, a big question for biology is what do species do? What, if any, are the functional properties of species?
Function • Anything that contributes to autonomy is functional (contributes to self-persistence), making selection possible when there is also variation among autonomous things. • Hypothesis: Function should be considered as the primary hierarchical property in biology
Structural versus functional decomposition • We can decompose biological entities (and other things with teleological properties) into both functional and structural parts. • These decompositions need not correspond • Consider decomposition of airplane versus bird into functions of lift and propulsion. • Although structural parts are restricted by function, and structure can tell a lot about function and contributions to function, I believe function is primary in biology (and in other teleological entities).
Example: Immune System • The immune system is not contained within specific parts of the body, but is spread out and uses parts of things like cells, cells them selves, and so on in very complex ways. • So there is no simple way to decompose the immune system structurally in a way the corresponds to typical structural decompositions of the body (organ, cell, cellules, etc.). • However, if it is regarded functionally, as a sort of organ, it is on a par with other organs like the heart, the skin, the nervous system, and so on. • But this suggests that the relevant aspects of these other organs for decomposition is not structural, but functional.
Some notes on reduction • Type reductions are ontological deflations – where there were many, we see that there are only few or perhaps one (Collier and Hooker). • Individual reductions are of wholes to parts. • Only possible completely if the wholes are aggregates. • These two types of reduction are sometimes confused (e.g., Kim) so that ontological deflation is taken to imply whole-part reduction. • This does not hold in general (consider fusion, spontaneous self-organization, emergence).
Thank you for your attention John Collier collierj@ukzn.ac.za http://Collier.ukzn.ac.za