Download

1 / 69
701 likes | 883 Views
REGULATOROWE RNA. Dawniej: RNA mRNA rRNA tRNA. Nie mo żna wyjaśnić złożoności organizmu ilością genów kodujących różne białka Człowiek 30 000 2% Drosophila melanogaster 13 000 13%

E N D
REGULATOROWE RNA
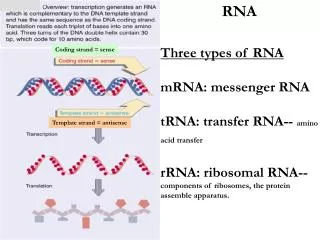
Dawniej: RNA mRNA rRNA tRNA
Nie można wyjaśnić złożoności organizmu ilością genów kodujących różne białka Człowiek 30 000 2% Drosophila melanogaster 13 000 13% Caenorhabditis elegans 17 000 27% Arabidopsis thaliana 25 000 50% Saccharomyces cerevisiae 6 000 57% Escherichia coli 4 000 >80% Ilość genów kodujących białka % DNA
ilość EST = 2 ilość ORF EST - expressed sequence tag ORF - open reading frame Stąd wniosek – duża część transkryptów nie koduje białek
RNA o funkcjach katalitycznych, często stanowią integralną część białek lub kompleksów białkowych takich jak: telomeraza, snRNP (zawierają małe jądrowe RNA), snoRNP (zawierają małe jąderkowe RNA), Rnaza P. Dziś: RNA Kodujące białka Niekodujące (mRNA) • konstytutywne • rRNA • tRNA • o aktywności • katalitycznej regulatorowe
Regulatorowe RNA • transkrypcja wielu z nich jest regulowana przez warunki środowiska czy stan zróżnicowania komórek lub jest tkankowo-specyficzna • wielkość: od 20 nukleotydów (nt) do >10 000 nt • stosunkowo mało konserwatywne (różnie, są wśród nich także konserwatywne) • występują i u Prokaryota i u Eukaryota • u eukariontów niektóre regulatorowe RNA są syntetyzowane przez polimerazę RNA II i podlegają takiej samej obróbce jak mRNA (mają czapeczkę, poli A i usuwane introny!); jedyna różnica – nie mają otwartej ramki odczytu.
INTRONY Introny przez wiele lat były postrzegane jedynie jako elementy zaangażowane w alternatywne składanie mRNA i jako rezerwuar materiału genetycznego do wykorzystania w dalszej ewolucji. Uważano, że introny są wycinane z pre-mRNA i natychmiast degradowane w jądrze komórkowym. • Co wiemy obecnie: • Niektóre introny (lub ich fragmenty) mogą mieć okres półtrwania porównywalny z mRNA • Niektóre introny (lub ich fragmenty) mogą być transportowane do cytoplazmy • Wniosek: • Niektóre introny pełnią jakąś funkcję!
Nie ma jednolitego nazewnictwa dla regulatorowych RNA: Bakteryjne regulatorowe RNA – small RNA Eukariotyczne regulatorowe RNA – noncoding RNA Stosowane są również terminy: Functional RNA (fRNA) Non-messenger RNA (nmsRNA) Wprowadza się również nazwy dla poszczególnych grup regulatorowych RNA np.: miRNA (mikroRNA), siRNA (small interfering RNA) itp.
Dwa główne mechanizmy regulacji: • Przez oddziaływanie z RNA o komplementarnej sekwencji nukleotydów • Przez oddziaływanie z białkami, których funkcjonowanie dotyczy kwasów nukleinowych (enzymy, czynniki transkrypcyjne). W tym przypadku ncRNA mogą działać jako inhibitory kompetycyjne.
Regulatorowe RNA, które oddziałują z RNA przez • parowanie zasad • kodowane na komplementarnej nici DNA • (cis-encoded RNA) – wykazują 100% komplementarności do jakiejś sekwencji regulowanego RNA • kodowane w innym miejscu genomu • (trans-encoded RNA) – znacznie niższa komplementarność
Dwa przykłady regulacji w komórkach bakteryjnych przez RNA kodowane przez plazmidowe DNA
Przykład 1: Mechanizm regulujący ilość plazmidów w komórce bakteryjnej. Plazmid pT181 (Staphylococcus) Białko RepC – inicjuje replikację plazmidu plazmidowe RNA I (85 nt) i RNA II (150 nt) są niestabilne i szybko ulegają degradacji
Gdy w komórce jest mało kopii plazmidu, to transkrypcji ulega odcinek obejmujący gen RepC
W miarę namnażania plazmidu przyrasta ilość RNA I i RNA II komplementarnych do odcinka poprzedzającego gen RepC. Oddziaływanie tych RNA z tworzonym mRNA tworzy strukturę powodującą terminację transkrypcji. Spada ilość RepC zahamowana replikacja plazmidu
Przykład 2: System uzależnienia od plazmidu (plasmid addiction system) Przeżywają bakterie zawierające plazmid, giną bakterie, które utraciły plazmid. (System hok-sok - plazmidR1 E. coli). hok – host killing sok – suppresor of killer Gdy w komórce jest plazmid, to toksyna nie jest produkowana bo translacja na hok mRNA jest blokowana przez sok RNA
Gdy bakteria traci plazmid, nie zachodzi transkrypcja hok i sok. Jednak sok RNA jest znacznie mniej stabilny niż hok mRNA. Zachodzi translacja, powstaje toksyna, która zabija bakterię.
Przykład regulacji w komórkach bakteryjnych przez RNA kodowane przez chromosomowe DNA
Białko GadX to czynnik transkrypcyjny regulujący odporność bakterii na kwaśne środowisko
Mechanizm odporności na kwaśne pH: GadX jest aktywatorem transkrypcji – pod jego wpływem wzrasta transkrypcja gadA i gadB; enzymów o aktywności dekarboksylazy glutaminianowej. Dekarboksylaza glutaminianowa przekształca glutaminian w kwas gamma-aminomasłowy z wykorzystaniem jednego protonu ze środowiska (z wnętrza komórki). GadC – antyporter – wymienia kwas g-aminomasłowy z komórki na glutaminian ze środowiska – umożliwia ciągły bieg reakcji.
Podobny typ regulacji stwierdzono także u ssaków Jeden z receptorów trijodotyroniny (T3) – TRa występuje w dwóch formach różniących się C-końcem (alternatywne składanie transkryptu). TRa1 wiąże hormon i odpowiednie miejsca regulatorowe na DNA. TRa2 wiąże się do DNA, ale nie aktywuje transkrypcji genów, bo nie wiąże ligandu. Jest inhibitorem kompetycyjnym dla aktywacji transkrypcji przez kompleks TRa1-T3. Odpowiedź komórki na T3 zależy więc od stosunku TRa1: TRa2. Wiadomo, że ten stosunek zależy od rodzaju tkanki i stopnia zróżnicowania komórek.
Sekwencja ostatniego egzonu TRa2 zachodzi na sekwencję ostatniego egzonu genu RevErb kodowanego na komplementarnej nici! Hastings et al. J.Biol.Chem. (2000), 275, 11507-13 RevErb – sierocy receptor jądrowy
Regulacja ekspresji czynnika transkrypcyjnego HIF1a HIF-1 (hypoxia inducible factorskłada się z 2 podjednostek HIF1a i b). Jego poziom w komórce rośnie w warunkach hipoksji. Stymuluje transkrypcję genów, których produkty mają zapobiegać hipoksji (np. VEGF). Poziom białka HIF1a podlega regulacji. W warunkach normoksji działa na niego hydroksylaza proliny co w rezultacie prowadzi do ubikwitynacji i degradacji. W warunkach hipoksji hydroksylaza jest nieaktywna i poziom białka HIF1a wzrasta.
Regulacja ekspresji czynnika transkrypcyjnego HIF1a poprzez wzrost stabilności transkryptu. W niektórych typach nowotworów zaobserwowano zwiększoną ekspresję aHIF1
miRNA - najlepiej poznane – stanowią jedną konkretną grupę regulatorowych RNA miRNA i siRNA - odpowiadają za proces RNAi miRNA – mikroRNA siRNA – short interfering RNA (albo small....)
RNAi – RNA interference Termin ten dotyczy takiego procesu, w który zaangażowane są małe RNA pochodzące z dwuniciowych prekursorów
Pierwszy raz zaobserwowano zjawisko interferencji RNA przez przypadek prowadząc prace nad uzyskaniem petunii o ciemnopurpurowym kolorze (lata 80 XXw). www.scq.ubc.ca www.bio-pro.de Plan: nadekspresja enzymu kodującego pigment Efekt: kwiatki albo całkiem białe albo z białymi paskami
Genetycy molekularni roślin wprowadzili termin posttranscriptional gene silencing(PTGS) • Jest to mechanizm umożliwiający roślinom walkę z wirusami. • Rośliny transgeniczne produkujące fragment białka wirusowego (a więc syntetyzujące fragment wirusowego RNA) były oporne na zakażenie tym wirusem. • Istotną kwestią jest obecność w komókach roślinnych enzymu RdRP – (RNA-dependent RNA polymerase). • Aktywność tego enzymu powoduje powstawanie dsRNA
miRNA zostały odkryte w trakcie badań nad rozwojem Caenorabditis elegans • 1993 - Pierwszym poznanym miRNA był lin-4. Badano mutacje, które prowadzą do nieprawidłowości w różnicowaniui rozwoju nicienia. Stwierdzono, że mutacja występuje w obszarze, który nie koduje żadnego białka. Ale ten fragment DNA koduje jakieś RNA. • Takie same defekty rozwojowe dawała mutacja w obrębie 3’UTR mRNA kodującego białko lin-14. • Okazało się, że lin-4 RNA zawiera sekwencję komplementarną do kilku sekwencji w 3’UTR lin-14.
Oddziaływanie lin-4 (miRNA) z 3’UTR mRNA lin-14 MicroRNAs: small RNAs with a bigrole in gene regulation.Lin He & Gregory J. Hannon, Nature Reviews Genetics 5, 522-531 (July 2004)
The Nobel Prize in Physiology or Medicine 2006 "for their discovery of RNA interference - gene silencing by double-stranded RNA" Andrew Z. Fire Craig C. Mello Stanford University School of Medicine Stanford, CA, USA University of Massachusetts Medical School Worcester, MA, USA Wg niektórych David Baulcombe (genetyka roślin, wirusy roślinne) też powinien był dzielić z nimi tę nagrodę
Obecnie uważa się, że ponad 30% transkryptów w komórkach ludzkich jest regulowana przez miRNA. miRNAs znaleziono także w prostych organizmach wielokomórkowych jak: gąbki czy jamochłony. Dane na temat wszystkich poznanych miRNA można znaleźć w bazie danych: http://microrna.sanger.ac.uk
Różne miRNA oddziałują z 3’UTR różnych mRNA. • Transkrypcja miRNA (pri-miRNA) podlega takiej • samej regulacji jak transkrypcja różnych mRNA. • Ekspresja wielu miRNA jest specyficzna tkankowo. • Zmiana ekspresji wielu miRNA jest obserwowana w • nowotworach.
Jak powstają miRNA? miRNA - 22 nt siRNA – dłuższe do 30 nukleotydów RISC – RNA induced silencing complex
Pri-miRNA Ok. 50% loci miRNA ssaków leży blisko loci innych miRNA. Ulegają one wówczas transkrypcji jako policistronowa jednostka transkrypcyjna. Inne miRNA mają własne, odrębne promotory. Niektóre loci miRNA znajdują się w regionach niekodujących, inne w intronach (ok. 40%) lub w obszarach które mogą być intronami lub egzonami w zależności od alternatywnego splajsingu.
RNaza III Drosha (występuje tylko u zwierząt) współdziała z innymi białkami U ssaków - z białkiem DGCR8 (DiGeorge syndrome critical region gene 8) U D. melanogaster i C.elegans – Pasha Drosha/DGCR8 oraz Drosha/Pasha tworzą tzw. kompleks mikroprocesorowy Obróbka pri-miRNA jest procesem kotranskrypcyjnym!!! Obróbka przez Drosha poprzedza splajsing, wycięcie pri-miRNA nie zaburza splajsingu. Przypuszczalnie Drosha działa na etapie tworzenia wczesnego splajsosomu (splajsosom już utworzony, intron jeszcze nie wycięty.
Która nić stanowi miRNA? http://www.sciencemag.org/feature/data/prizes/ge/2006/schwarz.dtl
RISC (RNA-induced silencing complex) Głównymi białkami RISC są białka argonautowe (ang. Argonauts). U ssaków tylko Ago2 ma aktywność nukleazową – która jest prawdopodobnie odpowiedzialna zarówno za degradację nici pasażerskiej jak i za degradację docelowego mRNA – aktywność Slicer Gdy w kompleksie nie ma Ago2 – za usunięcie nici pasażerskiej jest odpowiedzialna helikaza
Mechanizm walki z wirusami poprzez zjawisko RNAi i zaangażowanie siRNA jest charakterystyczne tylko dla niektórych organizmów: np. muszki owocowej i roślin. U C. elegans i u ssaków nie występuje ten proces. Zarówno C. elegans jak i ssaki mają tylko jeden enzym Dicer Muszka ma 2 – jeden współdziała z siRNA drugi z miRNA. A. thaliana – 4 enzymy Dicer, 10 białek Ago
Różnice pomiędzy miRNA a klasycznym siRNA • miRNA – endogenne; siRNA – egzogenne (wirusowe) • miRNA – prekursorem jest 70-80-nukleotydowa pętla • RNA, siRNA - prekursorem jest długie, dsRNA • miRNA – na ogół brak idealnej komplementarności; • siRNA – idealna komplementarność
Jak działają miRNA: • Oddziaływanie miRNA z 3’UTR może prowadzić do zahamowania translacji: • poprzez zahamowanie oddziaływania eIF4E (białko wiążące czapeczkę z eIF4G) • odłączenie rybosomów od mRNA • sekwestrację mRNA w ciałkach P • Oddziaływanie miRNA z 3’UTR może prowadzić do degradacji mRNA • Są również pojedyncze doniesienia, że miRNA mogą stymulować translację
miRNA U człowieka – okryto prawie 700 miRNA, analiza informatyczna wskazuje na możliwość istnienia blisko 1000 miRNA u człowieka Spośród 200, które były pozbane najpierw – 50% ma ortologi u kręgowców, a 25% u bezkręgowców 53 z nich jest unikalnych dla naczelnych!
Regulacja ekspresji miRNA Ekspresja większości miRNA jest specyficzna w zależności od etapu różnicowania i rozwoju oraz typu tkanki. Dokładna regulacja jest wymagana do prawidłowego funkcjonowania a deregulacja może prowadzić do różnych chorób w tym nowotworów. Kontrola na poziomie transkrypcji. Na przykład: czynniki transkrypcyjne zaangażowane w miogenezę: miogenina i MyoD1 (myoblast determination 1) indukują transkrypcję miR-1 i miR-133 podczas miogenezy.
Niektóre miRNA są pod transkrypcyjną kontrolą onkogenów lub supresorów nowotworzenia. Np. p53 aktywuje rodzinę miR-34 a MYC albo aktywuje albo powoduje represję wielu miRNA zaangażowanych w cykl komórkowy i apoptozę. Regulacja epigenetyczna– locus miR203 ulega metylacji DNA w komórkach chłoniaka wywodzącego się z komórek T ale nie w normalnych limfocytach T.
Regulacja ekspresji miRNA może mieć znaczenie przy rozwoju nowotworów Przykład: 3’UTR mRNA kodującego białko Ras ma kilka powtórzeń sekwencji LCS (Let-7 complementarity site). Oddziaływanie miRNA let-7 z 3’UTR mRNA Ras hamuje ekspresję Ras. W nowotworach płuc u człowieka obserwuje się wysoki poziom Ras i niski poziom let-7 miRNA.
miRNA są zaangażowane w regulację wielkości organizmów i tkanek. Nadekspresja genu bantam u D. melanogaster powodowała nadmierny wzrost tkanki; mutacja powodowała powstawanie mniejszych, ale proporcjonalnie zbudowanych muszek. Gen bantam koduje miRNA bantam Bantam stymuluje proliferację, hamuje apoptozę. Wiąże się do 3’ UTR proapopotycznego Hid www.rupert-fish.co.uk/poultry%20pics/old_engl...
Kłopoty z klasyfikacją Obecnie endogenne małe regulatorowe RNA dzieli się na: miRNA piRNA esiRNA
miRNA – 22 nt, obróbka Drosha i Dicer, wiążą się z podrodziną białek Ago (rodzina Ago). Potranskrypcyjna regulacja różnych mRNA. piRNA – 24-31, obróbka nie zależy od Dicer, występują najobficiej w komórkach zarodkowych, wiążą się z podrodziną białek Piwi (rodzina Ago), niektóre sązaangażowane w wyciszanie transpozonów (degradacja, metylacja). Nie występują u roślin. esiRNA – 21 n, głównie poznane u D. melanogaster; występują też w oocytach myszy i embrionalnych komórkach macierzystych (mysz). Pochodzą z długich dwuniciowych RNA. Obróbka – Dicer ale nie Drosha. Wycisznie transpozonów, potranskrypcyjna regulacja, .