Download

1 / 80
800 likes | 1.05k Views
Chapter11 DNA Replication and Synthesis.

E N D
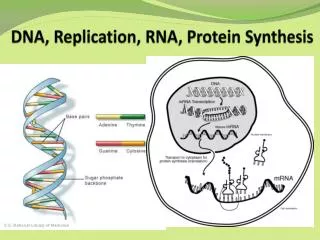
Chapter11DNA Replication and Synthesis Semiconservative replication of DNA makes genetics continuity between parental and progeny cells possible, as predicted by Watson-Crick model.Each strand of the parent helix serves as a template for its complement. DNA synthesis is a complex but orderly process orchestrated by a myriad of enzyme and other molecules. Together, they function with great fidelity to polymerize nucleotides into polynucleotide chains. Other enzymes interact with DNA, leading to genetic recombination.
Key topics in this chapter 11.1 The mode of DNA replication 11.2 synthesis of DNA in microorganisms 11.3 DNA synthesis : A model 11.4 A coherent mode of DNA synthesis 11.5 Genetic control of replication 11.6 Eukaryotic DNA synthesis 11.7 DNA replication ,telomeres and telomerase 11.8 DNA recombination
1. The model of DNA Replication DNA replication is semi-conservative, one strand serves as the template for the second strand. Furthermore, DNA replication only occurs at a specific step in the cell cycle. The following table describes the cell cycle for a hypothetical cell with a 24 hr cycle
Stage Activity Duration G1 Growth and increase in cell size 10 hr S DNA synthesis 8 hr G2 Post-DNA synthesis 5 hr M Mitosis 1 hr • DNA replication has two requirements that must be met: • DNA template • Free 3' -OH group
The Meselson-Stahl Experiment • In 1958, Matthew Meselson and Franklin Stahl published the results of an experiment providing strong evidence that cells use semiconservative replication to produce new DNA molecules. Escherichia coli cells were grown for many generations in a medium where I5NH4C1 (ammonium chloride) was the only nitrogen source. A "heavy" isotope of nitrogen, I5N contains one more neutron than the naturally occurring 14N isotope. Unlike "radioactive" isotopes, 15N is stable and thus does not decay (i.e., it is not radioactive).
After many generations, all nitrogen-containing molecules, including the nitrogenous bases of DNA, contained the heavier isotope in the E, coli cells. DNA containing I5N can be distinguished from '4N-containing DNA by the use of. sedimentation equilibrium centrifugation, in which centrifugation "forces" samples through a density gradient of a heavy metal salt such as cesium chloride.
The more dense 15N-DNA reaches equilibrium in the gradient at a point closer to the bottom (where the density is greater) than14N-DNA
Semiconservative Replication in Eukaryotes • In 1957, the year before the work of Meselson and his colleagues was published, J. Herbert Taylor, Philip Woods, and Walter Hughes presented evidence that semiconservative replication also occurs in eukaryotic organisms. They experimented with root tips of the broad bean Vicia faba, which are an excellent source of dividing cells. These researchers examined the chromosomes of these cells following replication of DNA. They monitored the replication process by labeling DNA with 3H-thymidine, a radioactive precursor of DNA, and then performing autoradiography.
Autoradiography is a cytological technique that pinpoints the location of an isotope in a cell. In this procedure, a photographic emulsion is placed over a section of cellular material (root tips in this experiment), and the preparation is stored in the dark. The slide is then developed, much as photographic film is processed. Because the radioisotope emits energy, the emulsion turns black at the approximate point of emission. The end result is the presence of dark spots or "grains" on the surface of the section, locating the newly synthesized DNA in the cell.
Origins, Forks, and Units of Replication • origin of replication. • Where along the chromosome is DNA replication initiated? • Is there only a single origin, or does DNA synthesis begin at more than one point? • Is a point of origin random or is it located at a specific region along the chromosome? • Second, once replication begins, does it proceed in a single direction or in both directions away from the origin? In other words, is replication unidirectional or bidirectional?
replication fork • Replicon the length of DNA that is replicated following one initiation event at a single origin is a unit called the,
In E. coli this specific region, called oriC, has been mapped along the chromosome. It consists of 245 base pairs, though only a small number are essential to the initiation of DNA synthesis. 3 9mers 4 11mers Since in bacteriophages and bacteria DNA synthesis originates at a single point, the entire chromosome constitutes one replicon. The presence of only a single origin is characteristic of bacteria, which have only one circular chromosome.
11.2 synthesis of DNA in microorganisms Proteins and enzymes of DNA Replication • DNA exists in the nucleus as a condensed, compact structure. To prepare DNA for replication, a series of proteins aid in the unwinding and separation of the double-stranded DNA molecule. These proteins are required because DNA must be single-stranded before replication can proceed.
DNA Helicases - These proteins bind to the double stranded DNA and stimulate the separation of the two strands. • DNA single-stranded binding proteins - These proteins bind to the DNA as a tetramer and stabilize the single-stranded structure that is generated by the action of the helicases. Replication is 100 times faster when these proteins are attached to the single-stranded DNA. • DNA Gyrase - This enzyme catalyzes the formation of negative supercoils that is thought to aid with the unwinding process. In addition to these proteins, several other enzymes are involved in bacterial DNA replication.
4. Primase - The requirement for a free 3' hydroxyl group is fulfilled by the RNA primers that are synthesized at the initiation sites by these enzymes. 5. DNA Ligase - Nicks occur in the developing molecule because the RNA primer is removed and synthesis proceeds in a discontinuous manner on the lagging strand. The final replication product does not have any nicks because DNA ligase forms a covalent phosphodiester linkage between 3'-hydroxyl and 5'-phosphate groups.
6. DNA Polymerase - DNA Polymerase I (Pol I) was the first enzyme discovered with polymerase activity, and it is the best characterized enzyme. Although this was the first enzyme to be discovered that had the required polymerase activities, it is not the primary enzyme involved with bacterial DNA replication. That enzyme is DNA Polymerase III (Pol III). Three activities are associated with DNA polymerase I; • 5' to 3' elongation (polymerase activity) • 3' to 5' exonuclease (proof-reading activity) • 5' to 3' exonuclease (repair activity) The second two activities of DNA Pol I are important for replication, but DNA Polymerase III (Pol III) is the enzyme that performs the 5'-3' polymerase function.
DNA Polymerases II and III • Although DNA synthesized under the direction of polymerase I demonstrated biological activity, a more serious reservation about the enzyme's true biological role was raised in 1969. Peter DeLucia and John Cairns reported the discovery of a mutant strain of E. coli that was deficient in polymerase I activity. The mutation was designated polA1. In the absence of the functional enzyme, this mutant strain of E. coli still duplicated its DNA and reproduced successfully! Other properties of the mutation led DeLucia and Cairns to conclude that in the absence of polymerase I, these cells are highly deficient in their ability to "repair" DNA.
For example, the mutant strain is highly sensitive to ultraviolet light and radiation, both of which damage DNA and are therefore mutagenic. Non mutant bacteria are able to repair a great deal of UV-induced damage. These observations led to two conclusions: • 1.At least one other enzyme that is responsible for replicating DNA in vivo is present in E. coli cells. • 2.DNA polymerase I may serve a secondary function in vivo. This function is now believed by Komberg and others to be critical to the fidelity of DNA synthesis,but this enzyme does not actually synthesize the entire complementary strand during replication.
Table 11.2 Properties of Bacterial DNA Pol I, II andIII Properties I II III Initiation - - - 5’-3’ Polymerization + + + 3’-5’ exonuclease activity + + + 5’-3’ exonuclease activity + - - Molecules of pol/cell 400 ? 15
Gene Function dnaA,I,P Initiation dnaB,C Helicase at oriC dnaE,N,Q,X,Z Subunits of DNA polymerase III dnaG Primase gyrA,B Subunits of gyrase lig Ligase oriC Origin of Replication polA DNA polymerase I polB DNA polymerase II rep Helicase ssb Single-stranded DNA binding proteins
11.3 DNA Synthesis : A General Model for DNA Replication • The DNA molecule is unwound and prepared for synthesis by the action of DNA gyrase, DNA helicase and the single-stranded DNA binding proteins. • A free 3‘-OH group is required for replication, but when the two chains separate no group of that nature exists. RNA primers are synthesized, and the free 3'OH of the primer is used to begin replication. • The replication fork moves in one direction, but DNA replication only goes in the 5' to 3' direction. This paradox is resolved by the use of Okazaki fragments.
Unwinding the DNA helix This region of the E. coii chromosome has been particularly well studied. Called oriC, it consists of 245 base pairs characterized by repeating sequences of 9 and 13 bases (called 9mers and 13mers). One particular protein (called DnaA because it is encoded by the gene dnaA) initiates unwinding of the helix. A number of subunits of the DnaA protein bind to each of several 9mers. This step is essential in facilitating the subsequent binding of DnaB and DnaC proteins that further open and destabilize the helix (Figure 11-9). Proteins such as these, which require the energy normally supplied by ATP hydrolysis to break hydrogen bonds and denature the double helix, are called helicases. Other proteins, called single-stranded binding proteins (SSBPs) stabilize
4.These are short, discontinuous replication products that are produced off the lagging strand. This is in comparison to the continuous strand that is made off the leading strand. 5.The final product does not have RNA stretches in it. These are removed by the 5' to 3' exonuclease action of Polymerase I. 6.The final product does not have any gaps in the DNA that result from the removal of the RNA primer. These are filled in by the action of DNA Polymerase I. 7. DNA polymerase does not have the ability to form the final bond. This is done by the enzyme DNA ligase.
Initiation of DNA synthesis Continuous and Discontuous DNA synthesis Concurrent synthesis on the leading and lagging strands Proofreading and error correction during DNA replication
11.6 Eukaryotic DNA Synthesis • Research shows that eukaryotic DNA is replicated in a manner similar to tha£ of bacteria.Eukaryotic polymerases have the same fundamental requirements for DNA synthesis as do bacterial systems; four deoxyribonucleoside triphosphates, a template, and a primer. However, because eukaryotic cells contain much more DNA per cell and because this DNA is complexed with proteins, eukaryotes face many problems not encountered by bacteria. As we might expect, these complications make the process of DNA synthesis much more complex in eukaryotes and more difficult to study. However, a great deal is now known about the process.
Multiple Replication Origins The most obvious difference between eukaryotic and prokaryotic DNA replication is that eukaryotic chromosomes contain multiple replication origins, in contrast to the single site that is part of the E. coli chromosome. Multiple Replication Origins Multiple Replication Origins The multiple origins are visible under the electron microscope (Figure 11-14).
They are essential if the entire genome of a typical eukaryote is to be replicated in a reasonable time. Recall that (1) eukaryotes have much greater amounts of DNA than bacteria (e.g., yeast has 4 times as much and Drosophila has 100 times as much DNA as E. coli); and (2) the rate of synthesis by eukaryotic DNA polymerase is much slower—only about 50 nucleotides per second, a rate 20 times less than the comparable bacterial enzyme. Under these conditions, single-origin replication of a typical eukaryotic genome would take up to one month to complete! However, replication is accomplished in as little as 3 minutes in some eukaryotic organisms. • Yeast 250-400 replicon, mammalian 2500
autonomously replicating sequences (ARSs). a unit of 11 base pairs, flanked by other short sequences involved in efficient initiation. DNA synthesis is restricted to the S phase of the eukaryotic cell cycle. Research has shown that the many origins are not all activated at once; instead clusters of 20-80 adjacent replicons are activated sequentially throughout the S phase until all DNA is replicated.
Eukaryotic DNA Polymerases • The most complex aspect of eukaryotic replication is the array of polymerases involved in directing DNA synthesis. Atotal of six different forms of the enzyme have been isolated and studied. For the polymerases to access DNA, the topology of the helix must first be modified. As synthesis is triggered at each origin site, the double strands are opened up in an A = T-rich region, which allows a helicase enzyme to enter that further unwinds the double-stranded DNA. before polymerases begin synthesis, histone proteins complexed to the DNA also must be stripped away or otherwise modified.
As DNA synthesis then proceeds, histones reassociate with the newly formed duplexes, reestablishing the characteristic nucleosome pattern Figure 11-15). In eukaryotes, the synthesis of new histone proteins is tightly coupled to DNA synthesis during the S phase of the cell cycle.
11.7 DMA Replication, Telomeres, and Telomerase • A final difference that exists between prokaryotic and eukaryotic DNA synthesis involves the nature of the chromosomes. Unlike the closed, circular DNA of bacteria and most bacteriophages, eukaryotic chromosomes are linear. During replication they face a special problem at the "ends" of these linear molecules, called the telomeres. While synthesis proceeds normally to the end of the leading strand, a difficulty arises on the lagging strand as the RNA primer is removed (Figure 11-16).
Normally, the newly created gap would be filled by adding a nucleotide to the existing 3'-OH group provided during discontinuous synthesis (to the right of gap b in Figure 11-16). However, there is no strand present to provide the 3 '-OH group because this is the end of the chromosome. As a result, each successive round of synthesis theoretically shortens the chromosome by the length of the RNA primer. Because this is such a significant problem, we can suppose, at least for some cells, that a molecular solution would have developed early in evolution and be shared by all
Centromeres and Telomeres Telomeres - the region of DNA at the end of linear eukaryotic chromosome; required for the replication and stability of the chromosome McClintock recognized several features about the ends of chromosomes • If two chromosomes were broken in a cell, the end of one could attach to the other and vice versa • She never observed was the attachment of the broken end to the end of an unbroken chromosome. • Thus the ends of broken chromosomes are sticky, whereas the normal end is not sticky, • This suggests the ends of chromosomes have unique features. • Usually, but not always, the telomeric DNA is heterochromatic and contains direct tandemly repeated sequences; the sequence TTAGGG is often the tandemly repeated sequence.
Species Repeat Sequence Arabidopsis TTTAGGG Human TTAGGG Oxytricha TTTTGGGG Slime Mold TAGGG Tetrahymena TTGGGG Trypanosome TAGGG Yeast (TG)1-3TG2-3 Telomere Repeat Sequences
Replication of Telomeres • Because DNA synthesis requires an RNA template (that provides the free 3'-OH group) to prime the synthesis, then a few bases at the end of a lagging strand would not be replicated during each round of replication • This would shorten the length of the chromosome after each division. But this is not seen. • Apparently telomerases, enzymes that replicate the terminal end of the chromosome exist. • These enzymes have a mode of action different than a normal DNA polymerase, but the end result is that sequences at the end of the chromosome do not go unreplicated during each replication round.
11.8 DNA Recombination • We conclude this chapter by returning to a topic discussed in Chapter 8—genetic recombination. There, we pointed out that the process of crossing over depends on breakage and rejoining of the DNA strands between homologs. Now that we have discussed the chemistry and replication of DNA.. it is appropriate to consider how recombination occurs at the molecular level. In general, the following information pertains to genetic exchange between any two homologous double-stranded DNA molecules, whether they be viral or bacterial chromosomes or eukaryotic homologs during Genetic exchange at equivalent positions along two chromosomes with substantial DNA sequence homology is referred to as general or homoloeous recombination.
Several models attempt to explain crossing over, and they all share certain common features. First, all are based on the initial proposals put forth independently by Robin Holliday and Harold L. K. Whitehouse in 1964. They also depend on the complementarity between DNA strands for their precision of exchange. Finally, each model relies on a series of enzymatic processes to accomplish genetic recombination. • One such model is shown in Figure 11-18. It begins with two paired DNA duplexes or homologs
Gene Conversion • A modification of the preceding model has helped us better understand a unique genetic phenomenon known as gene conversion(基因转变). • Initially found in yeast by Carl Lindegren and in Neurospora by Mary Mitchell, gene conversion is characterized by a genetic exchange ratio involving two closely linked genes that is nonreciprocal. If we cross two Neurospora strains each bearing a separate mutation (a+ X b+), a reciprocal recombination event between the genes yields spore pairs of the + + and ab genotypes. However, a nonreciprocal exchange yields one pair without the other.
Working with pyridoxine mutants, Mitchell observed several asci containing spore patterns displaying the ++ genotype, but not the reciprocal product (ab). Because the frequency of these events was higher than the predicted mutation rate and thus could not be accounted for by that phenomenon, they were called "gene conversions." They were so named because it appeared that one allele had somehow been "convened" to another during an event in which genetic exchange also occurred. Similar findings are apparent in the study of other fungi as well.
Gene conversion is now considered to be a consequence of the recombination process. One possible explanation interprets conversion as a mismatch of base pairs during heteroduplex formation, as shown in Figure 11—19. Mismatched regions of hybrid strands can be repaired by excising one
基因转变与异常分离 (遗传学 第8)p198基因转变Gene conversion及其分子机制 • 通常认为,重组总是交互的。 例1:脉孢菌:杂合体Aa,一个染色体把基因A交给它的同源染色体,同源染色体必然把基因a返回来叫给它。所以真菌中的一个位点(locus)上两个等位基因的分离应呈 2:2分离。 • 但是在脉孢菌中发现,有的子囊含有(3A+1a)或(1A+3a)的子囊孢子。 • Mitchell对这种现象进行了详细的分析。他的杂交实验中用的是吡哆醇合成的基因。
有两个突变位点,接近: 一个位点上:一个突变基因pdxp——突变株在吡哆醇培养基上生长,对pH敏感,变酸后则不用再添加了。 相邻的一个位点:也有突变基因,也是吡哆醇需要型——但对pH不敏感pdx +pdxp x pdx + 杂交 对585个子囊孢子培养鉴定。结果: 在4个子囊中有野生型孢子对——好象两者之间发生了重组 但是,与预期的重组结果不同——没有出现双突变型(pdxp pdx)孢子对(图12-13)。