Download

1 / 78
780 likes | 799 Views
Topic 3. Lectures 5 and 6: Examples of Indirect Evidence for Past Evolution. So, we introduced indirect evidence for past evolution of 6 kinds: 1) suboptimality of the phenotype of one or many modern species, 2) homology, or unforced similarity, between phenotypes of different

E N D
Topic 3. Lectures 5 and 6: Examples of Indirect Evidence for Past Evolution So, we introduced indirect evidence for past evolution of 6 kinds: 1) suboptimality of the phenotype of one or many modern species, 2) homology, or unforced similarity, between phenotypes of different modern species, or between parts of their genomes, 3) (unforced) hierarchical distributions of states of variable traits within sets of modern species, 4) patterns in geographical distributions of modern species, not explainable by their adaptations, 5) agreement between what we see and a simple evolutionary scenario, 6) agreement between what we see and a partial theory of Macroevolution. Now we will consider examples of evidence of each kind.
1) Suboptimality Remember, suboptimality may be easy-to-improve and hard-to-improve, and must not be caused by a recent change in the environment, in order to provide evidence for evolution. Perhaps, (almost) all features of all organisms are to some extent suboptimal, but this is usually impossible to show. Thus, we will only deal with the most salient examples. Let us consider suboptimality at all levels, starting from sequences.
Faulty design at the genomic level: factor VIII gene About 50% of cases of severe hemophilia A are caused by inversions, due to illegitimate recombination within the factor VIII gene. This recombination involves long, apparently functionless, repeats inside the gene and close to it. Factor VIII intron 1 and the model for inversion. (A) Bar shows factor VIII intron 1, flanked by exons (drawn to indicate the direction of transcription), and containing a repeated sequence 9b (shaded) flanked by unique sequences 9a and 9c. The line shows DNA outside the F8 gene with the repeated sequence as a shaded box. Arrowheads indicate orientation of repeated sequences. The large curved arrow indicates the folding required for homologous recombination. (B,C) Homologous recombination between the two 9b repeats proposed to explain origin of inversion. (D) Inversion resulting from recombination. Blood 99, 168-174, 2002.
Such faulty designs are very common. At a number of loci causing human Mendelian diseases, a particular kind of large-scale genomic rearrangement occurs recurrently due to some kind of repeats. Within the human population, variation in the number of copies of genes, also caused by recombination involving repeats, is pervasive and sometimes leads to sporadic cases of mental and other diseases. Theory predicts that if a certain genomic feature produces a lethal mutation with probability below 10-5, this feature may become fixed in the population with effective size below 105 (which is te case for humans). Thus, existence of faulty genomic features may also be viewed as a theory-based evidence for evolution, because, indvidually, such features never lead to high-frequency mutations.
A complex machinery inserts just one amino acid Selenocysteine (Sec) is inserted into selenoproteins in response to codon UGA with the help of the selenocysteine insertion sequence (SECIS) element. The number of selenoproteins in animals varies, with humans having 25 selenoproteins. However, in Caenorhabditis elegans and C. briggsae genomes thioredoxin reductase is the only Sec-containing protein. In contrast, we identified additional selenoproteins in other nematodes. Assuming that Sec insertion mechanisms are conserved between nematodes and other eukaryotes, the data suggest that nematode selenoproteomes were reduced during evolution, and that in an extreme reduction case Sec insertion systems decode only a single UGA codon. Nucleic Acids Research 33, 2227-2238 (2005).
Suboptimal development: transient hindlimbs Transient hindlimb buds in embryonic development of the spotted dolphin, Stenella attenuata. B: 24 days of gestation - well-developed early hindlimb (h) and forelimb (f) buds. C: 48 days of gestation - well-developed forelimb bud (f; note the digital primordia) and a regressing hindlimb bud (h). In adult spotted dolphins, hindlimbs are completely absent. Of course, this is also an example of homology. Evol. & Dev. 4, 445-458, 2002.
Transient tooth buds in baleen whale embryo's Baleen whales have no teeth, but on the upper jaw have a series of baleen plates, which are composed of cornified epithelium. However, some baleen whales have temporary tooth buds on both the upper and lower jaws during their fetal period. These buds generally develop to the bell stage with dentin, but never generate enamel. Both the neogenerating baleen plate germ and degenerating tooth bud coexist on the upper jaw in the middle fetal period of the minke whale. The tooth bud degenerates by odontoclasts and macrophages, which resembles degeneration of deciduous teeth. J. of Vet. Med. Sci. 61, 227-232, 1999.
Rudimentary pelvic bones in whales: suboptimality or only homology? Rudimentary pelvic bone in a pilot whale, Globicephala sp. (D) Dorsal view, anterior below, with the last rib shown. (E) Side view of the pelvic bones and vertebral column. Note the general size (25 cm long) and orientation of the pelvic bones and their lack of connection to the vertebral column. Evol. & Dev. 4, 445-458, 2002. It is hard to be 100% sure that these bones are functionless - but their similarity with corresponding bones of terrestrial mammals must be homologous. Darwin: rudimentary structures are often extremely variable! This pattern may be considered as a theory-based evidence, because theory predicts that phenotypes which are not controlled by strong selection must accumulate variation.
Vestigial wings in Kiwi The wings are covered by the plumage and strongly reduced, so that any function appears unlikely.
Rudimentary wings and nerves in flightless grasshoppers In a flightless species, Barytettix psolus, adults lack flight muscles. These muscles are preserved and innervated during nymphal life, but disappear in the adult. Yet, its nerve persists and, in the adult, contains axonal presynaptic specializations opposite inappropriate targets such as glia and basal lamina (bottom). Our findings indicate that selective muscle death during development is one mechanism underlying the reduction of the flight system of Barytettix through evolution. J. of Neurobiology 17 (6): 627 - 636 (2004). Apoptosis is a common mechanism of development in a wide variety of organisms.
Terrestrial reproduction of sea turtles A very plausible suboptimality: a leatherback sea turtle Dermochelys coriacea, fully adapted to living in water, lays eggs on a beach. Only a minority of the new hatchlings will make it back to water.
Suboptimality of reproduction: gynogenesis (or pseudogamy) Some asexuals depend on sperm of sexual males to trigger embryogenesis, a reproductive mode called gynogenesis. The genetic information of males is typically not used. Theory predicts that such mating complexes are short-lived and highly unstable.
Some examples of gynogenetic (pseudogamic) forms of life: Carassius auratus gibelio Poecilia formosa Ambystoma laterale Hypericum perforatum
2) Homology cagctcaccatggatgatgatatcaccgcgctcgtcattgacaacggctc |||||||||||||||||||||||| ||||||||||| | ||||||||||| cagctcaccatggatgatgatatcgccgcgctcgtcgtcgacaacggctc cggcatgtgcaaggccagcttcacgggcgacaatgccgcccgggcagtct |||||||||||||||| ||||| |||||||| ||||| ||||||| |||| cggcatgtgcaaggccggcttcgcgggcgacgatgccccccgggccgtct tcccctccatcgttgggcaccccaggcaccag------------------ ||||||||||||| |||| ||||||||||||| tcccctccatcgtggggcgccccaggcaccaggtaggggagctggctggg -------------------------------------------------- tggggcagccccgggagcgggcgggaggcaagggcgctttctctgcacag -------------------------------------------------- gagcctcccggtttccggggtgggggctgcgcccgtgctcagggcttctt ----------------ggcgtgatggtgggcatgggtcagaaggattcct |||||||||||||||||||||||||||||||||| gtcctttccttcccagggcgtgatggtgggcatgggtcagaaggattcct atgtgggcgacgaggcccagagcaagagaggcat |||||||||||||||||||||||||||||||||| atgtgggcgacgaggcccagagcaagagaggcat Remember: homology may exist between different organisms and parts of the same organism (but only homology between genome segments provide evidence for evolution in this case), may involve functionless and functional phenotypes (in the later case, it is necessary to show that the same function can be performed by other means). Homologies provide the most pervasive and important evidence for past evolution of life. Let us consider homologies at different levels, starting from sequences.
Homology at the sequence level is everywhere! 1) Shared pseudogenes - discussed already 2) Similar orders of genes, including cases when adjacent genes perform unrelated functions. An average length of a region of synteny between human and murine genomes is ~5,000,000 nucleotides (40 colinear genes). We detected 558,000 highly conserved, reciprocally unique landmarks within the mouse and human genomes, which can be joined into conserved syntenic segments. The figure shows a typical 510-kb segment of mouse chromosome 12 that shares common ancestry with a 600-kb section of human chromosome 14 is shown. Blue lines connect the reciprocal unique matches in the two genomes. In general, the landmarks in the mouse genome are more closely spaced, reflecting the 14% smaller overall genome size. Nature 420, 520-562, 2002.
3) In closely related species, the corresponding amino acids in the corresponding proteins are usually encoded by the same codon (in ~99% of cases for humans and chimpanzees), although different synonymous codons could be used. Pt ATG GCC CTG TGG ATG CGC CTC CTG CCC CTG CTG GTG CTG CTG GCC CTC TGG GGA CCT GAC ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| | | ||| ||| ||| ||| ||| ||| ||| ||| Hs ATG GCC CTG TGG ATG CGC CTC CTG CCC CTG CTG GCG CTG CTG GCC CTC TGG GGA CCT GAC Pt CCA GCC TCG GCC TTT GTG AAC CAA CAC CTG TGC GGC TCC CAC CTG GTG GAA GCT CTC TAC ||| ||| | ||| ||| ||| ||| ||| ||| ||| ||| ||| || ||| ||| ||| ||| ||| ||| ||| Hs CCA GCC GCA GCC TTT GTG AAC CAA CAC CTG TGC GGC TCA CAC CTG GTG GAA GCT CTC TAC Pt CTA GTG TGC GGG GAA CGA GGC TTC TTC TAC ACA CCC AAG ACC CGC CGG GAG GCA GAG GAC ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| Hs CTA GTG TGC GGG GAA CGA GGC TTC TTC TAC ACA CCC AAG ACC CGC CGG GAG GCA GAG GAC Pt CTG CAG GTG GGG CAG GTG GAG CTG GGC GGG GGC CCT GGT GCA GGC AGC CTG CAG CCC TTG ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| Hs CTG CAG GTG GGG CAG GTG GAG CTG GGC GGG GGC CCT GGT GCA GGC AGC CTG CAG CCC TTG Pt GCC CTG GAG GGG TCC CTG CAG AAG CGT GGTATC GTG GAA CAA TGC TGT ACC AGC ATC TGC ||| ||| ||| ||| ||| ||| ||| ||| ||| |||| ||| ||| ||| ||| ||| ||| ||| ||| ||| Hs GCC CTG GAG GGG TCC CTG CAG AAG CGT GGCATT GTG GAA CAA TGC TGT ACC AGC ATC TGC Pt TCC CTC TAC CAG CTG GAG AAC TAC TGC AAC TAG ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| ||| Hs TCC CTC TAC CAG CTG GAG AAC TAC TGC AAC TAG Alignment of coding sequences of chimpanzee and human preproinsulins. Different nonsynonymous codons are red, different synonymous codons are blue.
Massive genome degeneration in Mycobacterium leprae Leprosy, a chronic human disease, results from infection with the obligate intracellular pathogen Mycobacterium leprae. M. leprae has the longest doubling time of all known bacteria and has thwarted every effort at culture in the laboratory. Comparing the 3.27-megabase genome sequence of the leprosy bacillus with that of Mycobacterium tuberculosis (4.41 Mb) provides clear explanations for these properties and reveals an extreme case of reductive evolution. Less than half of the genome contains functional genes but pseudogenes, with intact counterparts in M. tuberculosis, abound. Gene deletion and decay have eliminated many important metabolic activities including siderophore production, part of the oxidative and most of the microaerophilic and anaerobic respiratory chains, and numerous catabolic systems and their regulatory circuits. Nature 409, 1007-1011, 2001. Similarity between M. tuberculosis genes and corresponding M. leprae pseudogenes is homologous.
Mycobacterium leprae genome From the outside: circles 1 and 2 (clockwise and anticlockwise) genes on the - and + strands, respectively; circles 3 and 4, pseudogenes; 5 and 6, M. leprae specific genes; 7, repeat sequences; 8, G+C content; 9, G/C bias (G+C)/(G-C). See http://www.sanger.ac.uk/Projects/M_leprae
Lost ability to synthesize vitamin C in humans Traces of past gene losses can be found in many genomes. For example, humans and other primate genomes miss a gene that encodes L-gulono-gamma-lactone oxidase in other mammals. Instead, our genome contains the pseudogene, which clearly is the degenerated remnant of exons 4-7 of this gene. This missing enzyme is responsible for our inability to synthesize vitamin C. (Biochim. Biophys. Acta 1472(1-2), 408-411, 1999). Functional L-gulono-gamma-lactone oxidase gene consists of 11 exons: LOCUS NW_001495470 24712 bp DNA linear CON 22-DEC-2006 DEFINITION Bos taurus chromosome 8 genomic contig, reference assembly, whole genome shotgun sequence. gene 1..24712 /gene="GULO" mRNA join(1..111,1089..1190,3182..3307,4565..4659,5027..5117, 7939..8124,16771..16878,19077..19226,20061..20225, 23085..23175,23932..24712) /gene="GULO" /product="L-gulono-gamma-lactone oxidase" CDS join(109..111,1089..1190,3182..3307,4565..4659,5027..5117, 7939..8124,16771..16878,19077..19226,20061..20225, 23085..23175,23932..24060)
In the alignment of bovine L-gulono-gamma-lactone oxidase mRNA with the corresponding segment of the human genome 4 exons highlighted by red show similarity: 1st: 23723 .. 23807 (85) 2nd: 350 .. 434 (85) GAGAAGAAGCAGGTGACCATGGAGGCTGGCATCCTCCTGGCCAACCTGCACCCACAGCTGGATGAGCATGGCCTGGCCCTGTCCA |||||||||||||||||| ||||||| ||||||||||| || |||||||||| |||||||| |||| |||||||| |||||| GAGAAGAAGCAGGTGACCGTGGAGGCCGGCATCCTCCTAGCTGACCTGCACCCCCAGCTGGACAAGCACGGCCTGGCTTTGTCCA 1st: 41070 .. 41153 (84) 2nd: 739 .. 822 (84) GAAGAAATCTGAGGACTTCTGCTTCCTCTGGTTCCCACACAGCGAGAATGTCAGTGCCATCCACCAGGACCACACCAGCAAGGC ||||||||| || ||||| |||| ||||||||||||||||||||||||||||||| ||| ||||||||||||||| |||| | GAAGAAATCCGAATACTTCCGCTTTCTCTGGTTCCCACACAGCGAGAATGTCAGTGTAATCTACCAGGACCACACCAACAAGCC 1st: 49402 .. 49538 (137) 2nd: 906 .. 1044 (139) TGCAGTGCCTTGTGGGCTGGATCAATGGCTTTTTGTTTCTGACTCCTGTT---TGCCTGCCAGGAGAACAGCAACCTCAGCCCCAAGATCTTCACCCACG ||| |||||||||||||||||||| ||||||| ||||| |||||||| || |||| ||| |||||||||||| ||||||||||||| ||| TGCCAGGCCTTGTGGGCTGGATCAATCGCTTTTT-CTTCTGGCTCCTGTTCAATGGGAAGAAGGAAAACTGCAACCTCAGCCACAAGATCTTCACCTACG TGTGCCACTTCAAGCAGCATGTCCAGCACTGGGCCATCCC ||||| ||||||||||||||||||| ||||||||||||| AGTGCCGCTTCAAGCAGCATGTCCAGGACTGGGCCATCCC 1st: 50376 .. 50543 (168) 2nd: 1045 .. 1213 (169) CAGAAAGAAGACCACGGAGGCCCTGCTGGAGCTGAAGGCCGTGCTGGAGGCCCACCCTGAGGTGGTGTCCCACTACCTGGTGGGGGTACGCTTCACCTG- |||| ||||||||| |||||||||||||||||||||||| |||||||||| |||| |||| ||| ||||||||| |||| |||||||||||| | CAGAGAGAAGACCAAAGAGGCCCTGCTGGAGCTGAAGGCCATGCTGGAGGCGAACCCCAAGGTAGTGGCCCACTACCCCGTGGAGGTACGCTTCACTCGC GAGGATGACATCCTACTGAGCCCCTGCTTCCAGTGGGACAGCCGCTACCTGAACATCAACCTGTACAGG | ||| |||||||| |||||||||||||||||| | |||||| ||||| ||||||||| | |||||||| GGGGACGACATCCTGCTGAGCCCCTGCTTCCAGCGCGACAGCTGCTACATGAACATCATCATGTACAGG Are we like this bear? Clearly, this gene loss may be suboptimal - only primates (and Guinea pigs!) can suffer from scurvy. However, this may not be a problem for our ancestors. Still, homology is obvious.
Alpha-globin pseudogene in the the white-blooded Antarctic icefishes The blood of Antarctic icefishes (family Channichthyidae, suborder Notothenioidei) is completely devoid of hemoglobin. Icefishes have developed compensatory adaptations that reduce oxygen demand and enhance oxygen transport. Oxygen delivery to tissues occurs by carrying the gas physically dissolved in the plasma. The genomes of three icefish species retain transcriptionally inactive alpha-globin-related DNA sequences, which are truncated variants of the alpha-globin gene of the red-blooded fish, containing part of intron 2, all of exon 3, and the 3'-untranslated region. The icefish genomes have no beta-globin genes. Gene 295, 185-191, 2002.
Universality of the genetic code Despite minor variations, the genetic code is essentially the same in all life. This genetic code may possess some advantageous features, but it could hardly represent the best of all possibilities. Instead, it is probably a "frozen accident".
Universality of proteins Even the most dissimilar forms of life, Archeae, Eubacteria, and Eukaryota, possess many similar proteins. Example: Multiple sequence alignment of P-loop kinases. The alignment shows three conserved sequence regions. Proteins from bacteria, archea, and eukaryota are included into this alignment. J. of Mol. Biol. 333, 781-815, 2003). Examination of the patterns of phyletic distribution of distinct families suggests that the Last Universal Common Ancestor of all modern life forms encoded 15-18 distinct alpha/beta ATPases and nucleotide-binding proteins of the HUP class.
Deep similarity of vertebrates at early stages of development Mammalian embryos possess branchial arches, similar and homologous to gills arches of fishes. http://www.millerandlevine.com/km/evol/embryos/Haeckel.html
Deep similarity of organisms that live under very different environments There are many sets of deeply similar species that still occupy very different ecological niches. Such similarities are probably homologous. Example: turtles (order Testudines), with ~300 modern species. All turtles have shells, which consist of carapace and plastron, and many other common traits. However, different turtles live in very dissimilar habitats. Eastern Painted Turtle Chrysemys picta picta Snapping Turtle Chelydra serpentina Green Turtle Cheloniamydas Spotted Turtle Clemmys guttata
Another example: birds Different birds are doing totally different things - many fly, but some do not - but still all of them possess many common (and homologous) molecular and morphological traits that define them as birds Such examples are countless.
Compatibility between different species Very many pairs of substantially different organisms can produce hybrids, of various degrees of viability and fecundity. An ability of two organisms to hybridize indicate their profound (and homologous) similarity. Mules are sterile, but viable, which is already something - try to assemble even a bad computer from a mixture of Mac and PC parts. Female hybrids produced by Panthera leo mothers and P. tigris fathers are usually fertile. Narcissus bulbicodium x romieuxii. Thousands of fully fertile hybrids between distinct forms of life are known.
An example of within-genome homology of two functional genes Within-genome homology can involve not only gene-pseudogene pairs, but also pairs of functional genes and proteins. Similarity is very likely to be unforced when proteins perform totally dissimilar functions. Alignment of amino acid sequences of delta-crystalline, a transparent, enzymatically inactive protein abundant in eye lens (top) and of enzyme argininosuccinate lyase (bottom), both of chicken, Gallus gallus. Within-genome homology is an evidence for Weak Claim for the species. MATEGDKLLGGRFVGSTDPIMEILSSSISTEQRLTEVDIQASMAYAKALEKASILTKTEL MA+EGDKL GGRF GSTDPIME+L+SSI+ +QRL+EVDIQ SMAYAKALEKA ILTKTEL MASEGDKLWGGRFSGSTDPIMEMLNSSIACDQRLSEVDIQGSMAYAKALEKAGILTKTEL EKILSGLEKISEESSKGVLVMTQSDEDIQTAIERRLKELIGDIAGKLQTGRSRNEQVLTD EKILSGLEKISEE SKGV V+ QSDEDI TA ERRLKELIGDIAGKL TGRSRN+QV+TD EKILSGLEKISEEWSKGVFVVKQSDEDIHTANERRLKELIGDIAGKLHTGRSRNDQVVTD LKLLLKSSTSVISTHLLQLIKTLVERAAIEIDIIMPGYTHLQKALPIRWSQFLLSHAVAL LKLLLKSS SVISTHLLQLIKTLVERAA EID+IMPGYTHLQKALPIRWSQFLLSHAVAL LKLLLKSSISVISTHLLQLIKTLVERAATEIDVIMPGYTHLQKALPIRWSQFLLSHAVAL TRDSERLGEVKKRITVLPLGSGALAGNPLEIDRELLRSELDMTSITLNSIDAISERDFVV RDSERLGEVKKR++VLPLGSGALAGNPLEIDRELLRSELD SI+LNS+DAISERDFVV IRDSERLGEVKKRMSVLPLGSGALAGNPLEIDRELLRSELDFASISLNSMDAISERDFVV ELISVATLLMIHLSKLAEDLIIFSTTEFGFVTLFDAYSTGSSLLPQKKNPDSLELIRSKA EL+SVATLLMIHLSKLAEDLIIFSTTEFGFVTL DAYSTGSSLLPQKKNPDSLELIRSKA ELLSVATLLMIHLSKLAEDLIIFSTTEFGFVTLSDAYSTGSSLLPQKKNPDSLELIRSKA GRVFGRLAAILMVLKGIPSTFSKDLQEDKEAVLDVVDTLTAVLQAATEVISTLQVNKENM GRVFGRLAA+LMVLKG+PST++KDLQEDKEAV DVVDTLTAVLQ AT VISTLQVNKENM GRVFGRLAAVLMVLKGLPSTYNKDLQEDKEAVFDVVDTLTAVLQVATGVISTLQVNKENM EKALTPELLSTDLALYLVRKGMPIRQAQTASGKAVHLAETKGITINNLTLEDLKSISPLF EKALTPELLSTDLALYLVRKGMP RQA ASGKAVHLAETKGI IN LTLEDLKSISPLF EKALTPELLSTDLALYLVRKGMPFRQAHVASGKAVHLAETKGIAINKLTLEDLKSISPLF ASDVSQVFSVVNSVEQYTAVGGTAKAA ASDVSQVF++VNSVEQYTAVGGTAK++ ASDVSQVFNIVNSVEQYTAVGGTAKSS
Homology of variation within different species Unforced similarity between different species may consist of homologous variation within each of them. Homologous variation is another evidence of deep (and probably unforced) similarity between species Example 1: locus white in Drosophila. Wild-type Drosophila melanogaster - wild-type D. simulans looks the same, despite 12% genetic differences. Loss-of-function allele of white locus in D. melanogaster - loss-of-function alleles at the corresponding locus in D. simulans - and in any other Drosophila - cause exactly the same phenotype.
Example 2: variation in taste sensitivity to phenylthiocarbamide in humans and chimpanzees. This is an intriguing story - contrary to the long-held belief, this variation could not be inherited from the common ancestor. Still, it demonstrates unforced similarity between humans and chimpanzees. It was reported over 65 years ago that chimpanzees, like humans, vary in taste sensitivity to the bitter compound phenylthiocarbamide (PTC). This was suggested to be the result of a shared balanced polymorphism. In humans, variable PTC sensitivity is controlled by the segregation of two common alleles at the TAS2R38 locus, which encode receptor variants with different ligand affinities. Here we show that PTC taste sensitivity in chimpanzees is also controlled by two common alleles of TAS2R38; however, neither of these alleles is shared with humans. Instead, a mutation of the initiation codon results in the use of an alternative downstream start codon and production of a truncated receptor variant that fails to respond to PTC in vitro. Therefore, although Fisher et al.'s observations were accurate, their explanation was wrong. Humans and chimpanzees share variable taste sensitivity to bitter compounds mediated by PTC receptor variants, but the molecular basis of this variation has arisen twice, independently, in the two species. Nature 440: 930-934 (2006).
J. of Bone and Joint Surgery 62, 508-510, 1980. Atavisms - a kind of homologous variation such that a rare variant within a species is more similar to other species (and presumably to ancestors) that the normal phenotype. An intermediate situation, really - variation is similar to norm.
Quiz: Describe in some detail a your own example of homology. Why, in this example, what we see today implies evolution in the past?
3) Hierarchies Among vertebrates, placenta is "nested" within bearing live young. Among insects, complete metamorphosis is nested within having wings. Remember: distributions of traits produced by exclusively divergent evolution must be hierarchical; often - but not always - we expect evolution to be divergent; only unforced hierarchies is an evidence for evolution. Hierarchical distributions of traits permeate all life - Darwin already knew this. Thus, there will be just a few examples here - we will encounter heirarchies again when phylogenetic reconstructions will be considered.
Poor hierarchies (only 2 combinations of states of variable traits). Example 1: Birds have feathers, no teeth, beak, no milk, erythrocytes with nuclei, right aortic arch. Mammals have hair, teeth, no beak, milk, erythrocytes without nuclei, left aortic arch. Example 2: Angiosperms have flowers, stamens with 2 pairs of pollen sacs; male gametophyte reduced to 3 cells, female gametophyte reduced to 7 cells with 8 nuclei, closed carpel enclosing the ovules, endosperm produced after "double fertilization". All other plants: none of the above. In both cases hierarchies are probably unforced: there is no obvious reason why an organism producing milk cannot have the left aortic arch.
Rich hierarchy (3 combinations of pairs of variable traits).
However, not all joint distributions of traits are hierarchical - often, there are a lot of conflicts between pairs of traits within a set of species. Evolutionary explanation of conflicts is homoplasy, repeated origins of the same trait state. One can expect traits to be prone to homoplasy if they i) evolve fast and ii) can accept only a small number of states Let us consider homoplasy at the sequence level. If we associate a trait with each nucleotide site, conflicts may be common, because there are only 4 possible states of each trait, and mutations that substitute individual nucleotides occur relatively often: species 1: ACAAG species 2: ACAGG species 3: ATAAG species 4: ATAGG However, we can also associate sequence-level traits with more substantial changes, such as deletions, insertions, and inversions. Inversion: AAACGAGGGAAA > AAACCCTCGAAA We can expect such traits to be less prone to homoplasy and, thus, to produce hierarchical joint distributions - and this is true.
The character matrix for 67 microinversions in 15 species of mammals. Each column represents an inversion-associated trait. Red and green cells represent inversions in opposite orientations, and gray cells represent missing data. There is not a single pair-wise conflict between traits here, so that their joint distribution is hierarchical. PNAS, 103, 19824-1982, 2006. Such data are also perfect for phylogenetic reconstructions.
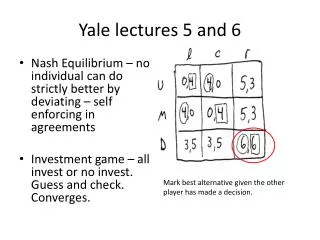
Congruent hierarchies If two kinds of organism are tightly connected to each other, we can expect them to evolve together, and produce congruent hierarchies, due to "cospeciation". Host-symbiont cospeciation in stinkbugs of the family Plataspidae, wherein a gut symbiotic bacterium is vertically transmitted via “symbiont capsule.” Removal of the symbionts resulted in retarded growth, mortality, and sterility of the insects. The host phylogeny perfectly agreed with the symbiont phylogeny, indicating strict host-symbiont cospeciation despite the extracellular association. PLoS Biology 10(4) 2006.
4) patterns in geographical ranges of similar species Unforced similarity of ranges of similar species is the simplest geographical pattern which provides evidence for Strong Claim. As usual, the challenge is to make sure that what we see cannot be explained just through adaptation to current conditions, without invoking evolution. This is obviously the case for shared absence of placentals in Australia, but may be questionable for shared absence of marsupials outside Australia (with some exceptions) - because invasions of marsupials are not common. When introduced, many placentals are doing great in Australia. Geographical evidence for evolution often rely on a particular evolutionary scenario - fortunately, it is not hard to figure out what geographical patterns to expect if evolution, indeed, occurred.
geographical pattern 1: local "flocks" of similar species are consistent with their evolution from the common ancestor under limited dispersal. A. Finches on the Galapagos islands There are 13 species of finches, which share a lot of traits that define them as finches, on the Galapagos islands, occupying a wide variety of ecological niches. http://www.rit.edu/~rhrsbi/GalapagosPages/DarwinFinch.html
B. Cichlids in Lake Malawi There are over 1000 species of cichlids in lake Malawi. These species use a wide variety of ecological niches. Similar niches may be occupied, in other places, by a wide variety of non-cichlid fishes. Obviously, this pattern is consistent with the local origin of these species from a common cichlid ancestor (or, perhaps, from several ancestors). Other African Great Lakes also contain cichlid species flocks of their own.
Cichlids: a family of mostly tropical fishes, characterized by a number of specific traits, including one nostril on each side (instead of two), and broken lateral line. Some other diagnostic anatomical traits of cichlids: 1. The loss of a major structural association between parts A2 and Aw of the adductor mandibulae muscle and the musculous insertion of a large ventral section of A2 onto the posterior border of the ascending process of the anguloarticular; 2. The presence of an extensive cartilaginous cap on the anterior margin of each second epibranchial bone; 3. The presence of an expanded head of each fourth epibranchial bone; 4. The presence of characteristically shaped and distributed micro-branchiospines on the gill arches; 5. The transversus dorsalis anterior muscle is subdivided into four distinct parts; 6. The stomach has an extendible blind pouch; 7. The stomach has a left hand exit to the anterior intestine and the first intestinal loop is on the left side; 8. The sagitta features an anterocaudal pseudocolliculum having a long and thick ventral part which is separated from the crista inferior by a long, deep and sharp furrow; 9. Short paired hypapophyses on the third and/or fourth vertebral centra (Kullander, 1998). Thus, a cichlid is not just a generic fish! Some charismatic examples of lake Malawi cichlids (many more can be found at http://malawicichlids.com and http://pick5.pick.uga.edu/):
Rhamphochromisesox. This and its congeners are known locally as "Mcheni" or "Ncheni". The species of Rhamphochromis are all pursuit predators, frequently feeding on the silvery, anchovylike minnow known as Usipa.
Dimidiochromis compressiceps, an ambush predator that eats whole small fish.
Buccochromis rhoadesii. Among the obliquely striped "haps" of Lake Malawi, this predatory, fish-eating species is most distinctive for its deep head (from top to bottom), with a very wide preorbital depth between the eye and the mouth.
Nimbochromis livingstonii is well known for its unusual predatory habits. It has been observed mimicking a dead fish, lying motionless on its side, on or partly covered by the substrate, and preying on small fish that come to pick at the "carcass".
Cheilochromis euchilus (adult, showing typical enlarged, fleshy lips, thought to form a seal over rock crevices enabling the fish to suck out aquatic insects).
Taeniolethrinops praeorbitalis lives close to sandy shores and plows into the substrate with its long snout, digging for the small invertebrates that are its food.