Download

1 / 44
440 likes | 923 Views
Cap. 13 Mappatura dei geni negli Eucarioti pp. 355-376. Sintesi 13. Non tutti gli alleli sono trasmessi indipendentemente, perché le posizioni dei geni (loci) sono associate sui cromosomi Scambi fra cromosomi omologhi alla meiosi provocano ricombinazione

E N D
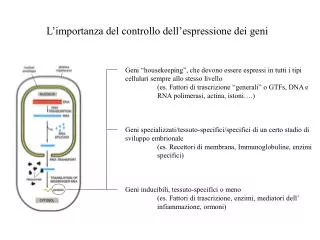
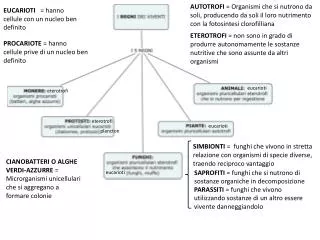
Sintesi 13 • Non tutti gli alleli sono trasmessi indipendentemente, perché le posizioni dei geni (loci) sono associate sui cromosomi • Scambi fra cromosomi omologhi alla meiosi provocano ricombinazione • Si può stimare la distanza fra due loci sulla base della frequenza di ricombinazione • Incrocio a tre punti
Trasmissione congiunta di alleli sul cromosoma X (Morgan 1911) E’ come un reincrocio
E quindi: Se prevalgono nella F2 i fenotipi parentali, la trasmissione dei due geni non è indipendente: geni associati sul cromosoma Se compaiono nella F2 fenotipi non parentali, l’associazione dei geni sul cromosoma non è completa, e gli alleli possono essere riassortiti da fenomeni di ricombinazione
Come indicare alleli associati a+ b a+ b a+ b a b+ ab a+ b+ Disposizione degli alleli: trans cis (repulsion) (coupling) La posizione occupata da un allele di un certo gene sul cromosoma si chiama locus
Cromosomi (e fenotipi) parentali e ricombinanti A b a B a b a b X Ab, aB = parentali AB, ab = ricombinanti
Il fenomeno citologico alla base della ricombinazione è il crossing-over
Esperimento di Barbara McClintock (1931) in Zea mais C: seme colorato, c: non colorato Wx: amido normale, wx: amido waxy Anche: carnation, Bar in Drosophila
Meccanismo della ricombinazione(Modello di Holliday,rottura ad elica singola)
Meccanismo della ricombinazione(Modello di Holliday,rottura ad elica singola) Eteroduplex: appaiamento temporaneo di eliche non complementari
Meccanismo della ricombinazione(Modello di Holliday,rottura ad elica singola)
L’intervento dell’endonucleasi può portare o non portare al crossing-over
Meccanismo della ricombinazione(Modello di Holliday, risoluzioni alternative dell’eteroduplex)
Meccanismo della ricombinazione(Modello di Holliday, conseguenze delle risoluzioni alternative)
Mappe genetiche Se il crossing over si verifica in punti casuali del cromosoma, la ricombinazione sarà più frequente fra loci distanti e meno fra loci vicini A B C bassa probabilità alta probabilità di crossing over di crossing over La percentuale di ricombinanti può essere utilizzata per quantificare la distanza fra loci associati: 1 Morgan (1 M) = 100% di ricombinazione 1 centiMorgan (1 cM) = 1% di ricombinazione
Il mezzo ideale per studiare associazione e ricombinazione è il reincrocio
Il mezzo ideale per studiare associazione e ricombinazione è il reincrocio Rapporti fenotipici attesi (diibrido): 1: 1: 1: 1 F(P) = F(NP) = 0,5 Se c’è associazione: F(P) > 0,5, F(NP)<0,5
Reincrocio Aa Bb X aa bb Frequenze fenotipiche: AB Ab aB ab TOT Osservate 78 122 118 82 400 Attese 100 100 100 100 400 Difetto Eccesso Difetto
Reincrocio Aa Bb X aa bb Frequenze fenotipiche: AB Ab aB ab TOT Osservate 78 122 118 82 400 Attese 100 100 100 100 400 I fenotipi Ab e aB sono presenti in frequenze superiori all’attesa Perciò Ab e aB sono i fenotipi parentali: A B a b A b a B La distanza fra locus A e locus B è d = NR / NT = =(78+82)/400 = 0,4 e cioè 40 cM
Ma attenzione: NR non può essere > ½ NT (in questo caso, si considerano i geni indipendenti) Per cui dMAX = 0,5.
Se tutti i cromosomi subiscono crossing-over alla meiosi, la frequenza di ricombinazione è del 50% La frequenza di ricombinazione è metà della frequenza di crossing over
Tre caratteri in Drosophila Vermillion (v) Crossveinless (cv) Cut (ct)
Starting with the v and cv loci, we see that the recombinants are of genotype v · cv and v+ · cv+ and that there are 45 + 40 + 89 + 94 = 268 of these recombinants. Of a total of 1448 flies, this number gives an RF of 18.5 percent. For the v and ct loci, the recombinants are v · ct and v+ · ct+. There are 89 + 94 + 3 + 5 = 191 of these recombinants among 1448 flies, so RF = 13.2 percent. L’incrocio può essere riscritto come v+ct cv / v ct+cv+ × v ct cv / v ct cv.
Interferenza Se i crossing over, CO, fossero indipendenti: P(doppio CO) = P(CO v-ct) x P(CO ct-cv) = = 0.132 x 0.064 = 0.0084 Doppi crossing over attesi, DCA = 1448 x 0.0084 = 12 Doppi crossing over osservati, DCO = 8 cc = coeff. di coincidenza = DCO/DCA = 0.67 I = interferenza = 1 – cc = 0.33
Macchie gemelle in Drosophila (Stern) In femmine y+ sn / y sn+
Macchie gemelle Frequenza troppo alta per essere dovute a doppia mutazione
Associazione fra DNA e malattie (broad genome scans) pazienti controlli
Caccia ai geni-malattia attraverso studi di associazione: associazione con la schizofrenia bipolare Segurado et al. 2003, Am. J. Hum. Genet.
La caccia ai geni-malattia attraverso studi di associazione ha dato risultati poco esaltanti Diabete di tipo 2: Associazioni (confermate e non confermate) con regioni dei cromosomi 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 14, 15, 16, 17, 20, 21 e X Alzheimer: 1, 2, 5, 6, 9, 10, 12, 13, 14, 19, 20, 21 e X Cancro alla prostata: 1, 2, 5, 6, 7, 8, 9, 10, 11, 16, 17, 18, 19, 20, 22, X Eterogeneità genetica, problemi con le popolazioni
Se la popolazione è strutturata, e la malattia è tipica di un certo gruppo, si troverà associazione per tutte le regioni di DNA in cui i gruppi sono diversi pazienti controlli (prevalentemente europei) (prevalentemente non europei)
Scelta sessuale Wedekind et al. Proc. Royal Soc. London 1995
Schema dell’esperimento • 49 femmine, 44 maschi. Tipizzati per gli antigeni HLA-A, -B, e –DR • I maschi indossano una T-shirt per due giorni • Per ogni femmina, 3 frammenti di T-shirt da maschio simile, 3 da maschio il cui HLA è dissimile (294 test) • Assegnati punteggi di intensità, piacevolezza e sex-appeal, da 0 a 10
Risultati Piacevolezza P=0.040 Ricorda un partner P=0.038 5 5 ≈ ≠ ≈ ≠ Conclusioni Anche al giorno d’oggi, la scelta del partner è associata ad alleli di loci HLA, o di loci che occupano posizioni vicine sui cromosomi.
Meccanismo della ricombinazione(Modello di Holliday, rottura nella doppia elica)NB: nel libro è presentato il modello con rottura di elica singola Eteroduplex: appaiamento temporaneo di eliche non complementari