Download

1 / 70
810 likes | 1.9k Views
Fatty Acid Synthesis. Lecture 16 Modified from internet sources, journals and books. Fatty Acid Synthesis. Prediction: the pathway for the synthesis of fatty acids would be the reversal of the oxidation pathway

E N D
Fatty Acid Synthesis Lecture 16 Modified from internet sources, journals and books
Fatty Acid Synthesis • Prediction: the pathway for the synthesis of fatty acids would be the reversal of the oxidation pathway • this would not allow distinct regulation of the two pathways to occur even given the fact that the pathways are separated within different cellular compartments • pathway for fatty acid synthesis occurs in the cytoplasm (oxidation occurs in the mitochondria) • the essential chemistry of the two processes reversals of each other
continued • oxidation and synthesis of fats utilize an activated two carbon intermediate acetyl-CoA • acetyl-CoA in fat synthesis exists temporarily bound to the enzyme complex as malonyl-CoA • synthesis of malonyl-CoA the first committed step of fatty acid synthesis • the enzyme that catalyzes this reaction acetyl-CoA carboxylase (ACC) = the major site of regulation of fatty acid synthesis
The rate of fatty acid synthesis • controlled by the equilibrium between monomeric ACC and polymeric ACC • activity of ACC requires polymerization the conformational change is enhanced by citrate and inhibited by long-chain fatty acids • ACC is also controlled through hormone mediated phosphorylation (see below). • The acetyl-CoA and malonyl-CoA are transferred to ACP (acetyl-CoA phosphatase) by the action of acetyl-CoA transacylase and malonyl-CoA transacylase, respectively
continued • attachment of these carbon atoms to ACP allows them to enter the fatty acid synthesis cycle. • The synthesis of fatty acids from acetyl-CoA and malonyl-CoA carried out by fatty acid synthase (FAS)
continued • All of the reactions of fatty acid synthesis are carried out by the multiple enzymatic activities of FAS (fatty acid synthase) • like fat oxidation fat synthesis involves 4 enzymatic activities: • β-keto-ACP synthase, β-keto-ACP reductase, 3-OH acyl-ACP dehydratase and enoyl-CoA reductase (the two reduction reactions require NADPH oxidation to NADP+) • the primary fatty acid synthesized by FAS is palmitate; then released from the enzyme and can then undergo separate elongation and/or unsaturation to yield other fatty acid molecules
Origin of Cytoplasmic Acetyl-CoA • Acetyl-CoA generated in the mitochondria primarily from two sources: • the pyruvate dehydrogenase (PDH) reaction • fatty acid oxidation • these acetyl units to be utilized for fatty acid synthesis they must be present in the cytoplasm • shift from fatty acid oxidation and glycolytic oxidation occurs when the need for energy diminishes • This results in reduced oxidation of acetyl-CoA in the TCA cycle and the oxidative phosphorylation pathway • Under these conditions the mitochondrial acetyl units can be stored as fat for future energy demands
continued • Acetyl-CoA enters the cytoplasm in the form of citrate via the tricarboxylate transport system • In the cytoplasm citrate is converted to oxaloacetate and acetyl-CoA (by the ATP driven ATP-citrate lyase reaction) • resultant oxaloacetate is converted to malate by malate dehydrogenase (MDH)
continued • The malate produced by this pathway can undergo oxidative decarboxylation by malic enzyme • co-enzyme for this reaction is NADP+ generating NADPH • advantage of this series of reactions for converting mitochondrial acetyl-CoA into cytoplasmic acetyl-CoA the NADPH produced by the malic enzyme reaction can be a major source of reducing co-factor for the fatty acid synthase activities
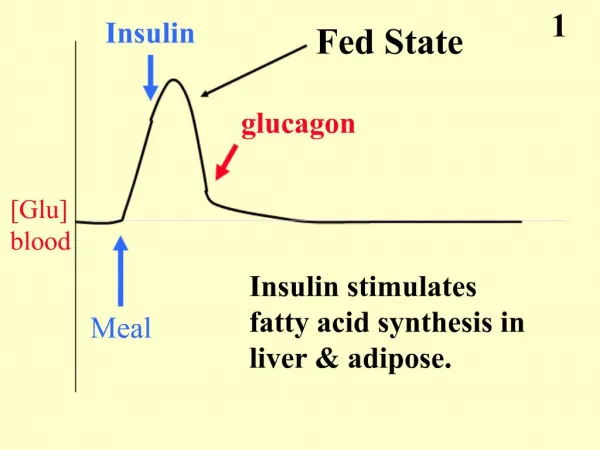
Regulation of Fatty Acid Metabolism • must consider the global organismal energy requirements in order to effectively understand how the synthesis and degradation of fats (and also carbohydrates) needs to be exquisitely regulated • blood is the carrier of triacylglycerols in the form of VLDLs and chylomicrons, fatty acids bound to albumin, amino acids, lactate, ketone bodies and glucose • The pancreas is the primary organ involved in sensing the organisms dietary and energetic states via glucose concentrations in the blood
continued • The regulation of fat metabolism occurs via distinct mechanisms: • short term regulation regulation effected by events such as substrate availability, allosteric effectors and/or enzyme modification • ACC (acetyl-CoA carboxylase) the rate-limiting (committed) step in fatty acid synthesis
continued • two major isoforms of ACC in mammalian tissues: • ACC1 and ACC2 • ACC1 is strictly cytosolic and is enriched in liver, adipose tissue and lactating mammary tissue • ACC2 originally discovered in rat heart but is also expressed in liver and skeletal muscle • Both isoforms of ACC allosterically activated by citrate and inhibited by palmitoyl-CoA and other short- and long-chain fatty acyl-CoAs
continued • Citrate triggers the polymerization of ACC1 which leads to significant increases in its activity • ACC2 does not undergo significant polymerization (presumably due to its mitochondrial association), is allosterically activated by citrate • Glutamate and other dicarboxylic acids can also allosterically activate both ACC isoforms
continued • ACC activity can also be affected by phosphorylation • Glucagon-stimulation increases in cAMP and subsequently increasing PKA activity also lead to phosphorylation of ACC and ACC2 • This insulin-mediated effect has not been observed in hepatocytes or adipose tissues cells • Activation of α-adrenergic receptors in liver and skeletal muscle cells inhibits ACC activity as a result of phosphorylation (undetermined kinase)
continued • Control of a given pathways' regulatory enzymes can also occur by alteration of enzyme synthesis and turn-over rates these changes are long term regulatory effects • Insulin stimulates ACC and FAS synthesis, whereas, starvation leads to decreased synthesis of these enzymes • Adipose tissue lipoprotein lipase levels also are increased by insulin and decreased by starvation
continued • in contrast to the effects of insulin and starvation on adipose tissue their effects on heart lipoprotein lipase are just the inverse • this allows the heart to absorb any available fatty acids in the blood in order to oxidize them for energy production • Adipose tissue contains hormone-sensitive lipase (HSL), that is activated by PKA-dependent phosphorylation leading to increased fatty acid release to the blood
continued • In the liver the net result of activation of HSL (due to increased acetyl-CoA levels) is the production of ketone bodies • This would occur under conditions where insufficient carbohydrate stores and gluconeogenic precursors were available in liver for increased glucose production • Insulin has the opposite effect to glucagon and epi leading to increased glycogen and triacylglyceride synthesis • One of the many effects of insulin to lower cAMP levels which leads to increased dephosphorylation through the enhanced activity of protein phosphatases
ChREBP: Master Lipid Regulator in the Liver • ChREBP = helix-loop-helix/leucine zipper (bHLH/LZ) transcription factor, carbohydrate-responsive element-binding protein has emerged as a central regulator of lipid synthesis in liver • ChREBP identified as a major glucose-responsive transcription factor and it is required for glucose-induced expression of the hepatic isozyme of the glycolytic enzyme pyruvate kinase (identified as L-PK) • ChREBP acts to induce lipogenic genes such as acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS)
continued • Expression of the ChREBP gene induced in the liver in response to increased glucose uptake • Under conditions of low (basal) glucose concentration ChREBP is phosphorylated and resides in the cytosol • An emerging model of the role of ChREBP in overall glucose and lipid metabolism indicates it a master regulator of glucose-mediated lipid homeostasis not only in the liver but also in adipose tissue
Elongation and Desaturation • The fatty acid product released from FAS is palmitate (a 16:0 fatty acid, i.e. 16 carbons and no sites of unsaturation) • Elongation and unsaturation of fatty acids occurs in both the mitochondria and endoplasmic reticulum • The predominant site of these processes the ER membranes • Elongation involves condensation of acyl-CoA groups with malonyl-CoA • resultant product two carbons longer (CO2 is released from malonyl-CoA as in the FAS reaction) which undergoes reduction, dehydration and reduction yielding a saturated fatty acid • Mitochondrial elongation involves acetyl-CoA units and is a reversal of oxidation
continued • Desaturation occurs in the ER membranes • involves 4 broad specificity fatty acyl-CoA desaturases (non-heme iron containing enzymes) • These enzymes introduce unsaturation at C4, C5, C6 or C9 • electrons transferred from the oxidized fatty acids during desaturation are transferred from the desaturases to cytochrome b5 and then NADH-cytochrome b5 reductase • These electrons are un-coupled from mitochondrial oxidative-phosphorylation and do not yield ATP
Since these enzymes cannot introduce sites of unsaturation beyond C9 they cannot synthesize either linoleate (18:2Δ9,12) or linolenate (18:3Δ9,12,15) • These fatty acids must be acquired from the diet referred to as essential fatty acids • Linoleic especially important in that it is required for the synthesis of arachidonic acid • arachindonate a precursor for the eicosanoids (the prostaglandins and thromboxanes)
continued • role of fatty acids in eicosanoid synthesis that leads to poor growth, wound healing and dermatitis in persons on fat free diets • linoleic acid a constituent of epidermal cell sphingolipids that function as the skins water permeability barrier
Synthesis of Triglycerides • Fatty acids stored for future use as triacylglycerols in all cells, but primarily in adipocytes of adipose tissue • fatty acids present in triacylglycerols predominantly saturated • major building block for the synthesis of triacylglycerols, in tissues other than adipose tissue, = glycerol • Adipocytes lack glycerol kinase dihydroxyacetone phosphate (DHAP), produced during glycolysis, is the precursor for triacylglycerol synthesis in adipose tissue • adipoctes must have glucose to oxidize in order to store fatty acids in the form of triacylglycerols
continued • The glycerol backbone of triacylglycerols activated by phosphorylation at the C-3 position by glycerol kinase • The fatty acids incorporated into triacylglycerols activated to acyl-CoAs through the action of acyl-CoA synthetases • Two molecules of acyl-CoA esterified to glycerol-3-phosphate to yield 1,2-diacylglycerol phosphate (commonly identified as phosphatidic acid).
continued • The phosphate is then removed to yield 1,2-diacylglycerol, the substrate for addition of the third fatty acid • Intestinal monoacylglycerols, derived from the hydrolysis of dietary fats, can also serve as substrates for the synthesis of 1,2-diacylglycerols
Phospholipid Structures • Phospholipids synthesized by esterification of an alcohol to the phosphate of phosphatidic acid (1,2-diacylglycerol 3-phosphate) • Most phospholipids a saturated fatty acid on C-1 and an unsaturated fatty acid on C-2 of the glycerol backbone • The most commonly added alcohols = serine, ethanolamine and choline • The major classifications of phospholipids are:
PC • This class of phospholipids also called the lecithins • At physiological pH phosphatidylcholines are neutral • contain primarily palmitic or stearic acid at carbon 1 and primarily oleic, linoleic or linolenic acid at carbon 2 • lecithin dipalmitoyllecithin a component of lung or pulmonary surfactant • the major (80%) phospholipid found in the extracellular lipid layer lining the pulmonary alveoli
PE • These molecules are neutral at physiological pH • contain primarily palmitic or stearic acid on carbon 1 and a long chain unsaturated fatty acid (e.g. 18:2, 20:4 and 22:6) on carbon 2
PS • composed of fatty acids similar to the phosphatidyl-ethanol-amines • PE is in the lipid bilayer of the a membrane
PI • contain almost exclusively stearic acid at carbon 1 and arachidonic acid at carbon 2 • molecules exist in membranes with various levels of phosphate esterified to the hydroxyls of the inositol • Molecules with phosphorylated inositol polyphosphoinositides • polyphosphoinositides important intracellular transducers of signals emanating from the plasma membrane
continued • One polyphosphoinositide (phosphatidylinositol 4,5-bisphosphate, PIP2) a critically important membrane phospholipid involved in the transmission of signals for cell growth and differentiation from outside the cell to inside
PG • Phosphatidylglycerols found in high concentration in mitochondrial membranes and as components of pulmonary surfactant • Phosphatidylglycerol a precursor for the synthesis of cardiolipin (important component of the inner mitochondrial membrane, where it constitutes about 20% of the total lipid) • vital role of PG serve as the precursor for the synthesis of diphosphatidylglycerols (DPGs)
DPG • These molecules very acidic • primarily in the inner mitochondrial membrane and also as components of pulmonary surfactant
continued • The fatty acid distribution at the C-1 and C-2 positions of glycerol within phospholipids is continually in flux • phospholipid degradation and the continuous phospholipid remodeling that occurs while these molecules are in membranes (= highly dynamic systems) • Phospholipid degradation results from the action of phospholipases • various phospholipases exhibiting substrate specificities for different positions in phospholipids • remodeling of acyl groups in phospholipids = the result of the action of phospholipase A1 (PLA1) and phospholipase A2 (PLA2)
continued • products of these phospholipases called lysophospholipids and can be substrates for acyl transferases utilizing different acyl-CoA groups • PLA2 an important enzyme, whose activity is responsible for the release of arachidonic acid from the C-2 position of membrane phospholipids • released arachidonate a substrate for the synthesis of the eicosanoids • there is not just a single PLA2 enzyme; At least 19 enzymes have been identified with PLA2 activity involved in numerous processes including modification of eicosanoid generation, host defense, and inflammation
The cytosolic PLA2 family (cPLA2) essential component of the initiation of arachidonic acid metabolism • the sPLA2 enzymes tightly regulated by Ca2+ and by phosphorylation
Plasmalogens • Plasmalogens are glycerol ether phospholipids • Three major classes of plasmalogens have been identified: • choline, ethanolamine and serine plasmalogens • Ethanolamine plasmalogen prevalent in myelin • Choline plasmalogen abundant in cardiac tissue. • One choline (1-O-1'-enyl-2-acetyl-sn-glycero-3-phosphocholine) identified as an extremely powerful biological mediator is called platelet activating factor= PAF
continued • PAF functions as: • a mediator of hypersensitivity, acute inflammatory reactions and anaphylactic shock • PAF is synthesized in response to the formation of antigen-IgE complexes on the surfaces of basophils, neutrophils, eosinophils, macrophages and monocytes • synthesis and release of PAF from cells leads to platelet aggregation and the release of serotonin from platelets • PAF also produces responses in liver, heart, smooth muscle, and uterine and lung tissues