Download
1 / 24
260 likes | 531 Views
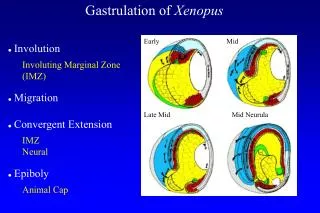
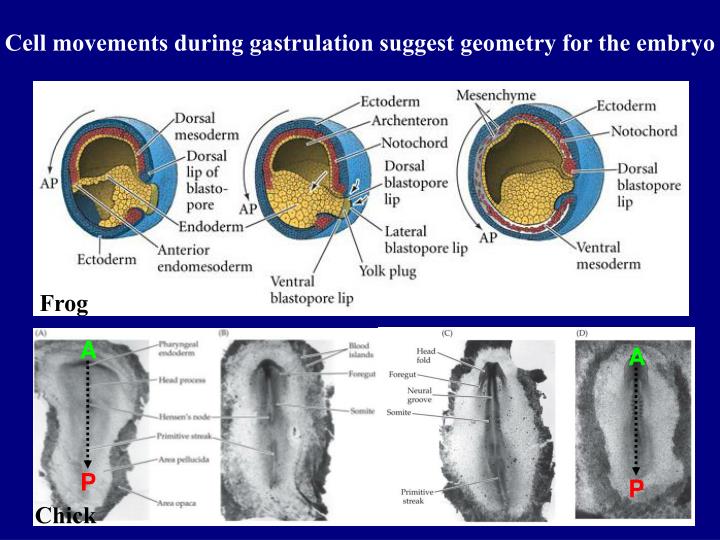
Frog. A. A. P. P. Chick. Cell movements during gastrulation suggest geometry for the embryo. Chordin. Chordin. Chordin. Dorsal lip of blastopore. Frog. Chick. Gene expression, particularly in mesoderm, and local signaling distinguish

E N D
Frog A A P P Chick Cell movements during gastrulation suggest geometry for the embryo
Chordin Chordin Chordin Dorsal lip of blastopore Frog Chick Gene expression, particularly in mesoderm, and local signaling distinguish A-P axis from early gastrulation onward in register with cell movements …and antagonistic signaling establishes further local and axial distinctions
head tail The geometry of cell movements during gastrulation in the mouse is distinct, but the results are the same…
antagonist signal TF target Noggin TF target Germ layers are divided into anterior (head) and posterior (tail) territories by antagonistic signaling pathways that result in local expression of transcription factor targets
noggin Bmps chordin Dkk SFRPs Wnts Lefty Cerberus Nodal Essential antagonist/agonist pairs of signals, from distinct local sources establish A-P patterning in the germ layers Anteriorizing Posteriorizing The localization of antagonists in the anterior region suggests that “head” is a default, and “tail” must be actively constructed
+/+ Nog:Chord-/- Chord-/- Loss of Bmp antagonist function disrupts head development
Loss of Wnt antagonism via Dkk causes head to be transformed into posterior tissue
Nodal signaling uses pathways similar to that for Bmp: both are members of the Tgfb superfamily of signals
Hex is a marker for visceral endoderm Loss of Nodal antagonism causes expansion of visceral endoderm, normally restricted to posterior of embryo
Heads or Tails: The balance of Bmp, Wnt and Nodal signaling decides!
There is more to posterior development than making a tail: posterior regionalization via the Hox genes Colinear expression relies on temporally controlled chromatin remodeling in a 3’to 5’ direction Paralogues on different chromsomes: 3’=anterior, 5’=posterior
most posteriorly restricted in all 3 germ layers Is there a terminal posterior identity: ParaHox genes and establishing the end of the “tail”
Cyp26 RA degrading enzyme Cyps Hoxb1 RA Raldh2 RA synthesizing enzyme Head/Tail antagonism holds for RA signaling Posterior signals include Wnts, and Fgfs: establish graded gene expression in concert with RA Regulation of Hox/ParaHox expression reflects antagonist/agonist signaling
Hox in the head: maintaining posterior segmentation in anteriorized territory
ParaHox and Hox genes are central regulators of A-P identity
1.Conditional mutation of Cdx2 in the post-gastrula endoderm causes posterior gut dismorphogenesis
goblet cells (alcian blue) 2a.Primary differentiated cellular characteristics of intestinal epithelium are absent in posterior gut of Cdx2 conditional mutant enterocytes: alkaline phosphatase
2b. Cellular architecture is disrupted, and cell proliferation is altered in posterior gut of Cdx2 conditional mutant
3. The dysmorphogenic posterior gut has been “anteriorized” to resemble esophageal epithleium by loss of Cdx2 in post-gastrula endoderm.
4. Anteriorization of gut epithelium to esophageal epithelium is accompanied by shifted expression (spatial or temporal) of anterior endoderm genes, including Hox cluster
5. Shift of expression of Wnts, transcriptional regulators, and down stream targets (all evidence of M-E signaling in which Wnt10a, 3a are available from M and act on E) in anterior gut, and in posterior mutant gut