Download
1 / 1
20 likes | 163 Views
The neural basis of unconscious orthographic priming Joseph. T. Devlin, Sharon Geva, Hannah Devlin, Mark W. Woolrich Centre for Functional Magnetic Resonance Imaging of the Brain University of Oxford, Oxford, U. K. C35. Email: devlin@fmrib.ox.ac.uk. Predictions.

E N D
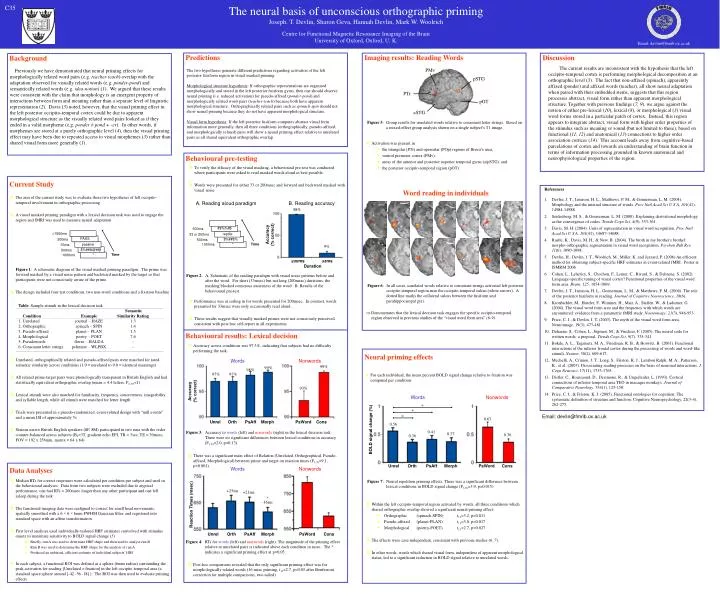
The neural basis of unconscious orthographic priming Joseph. T. Devlin, Sharon Geva, Hannah Devlin, Mark W. Woolrich Centre for Functional Magnetic Resonance Imaging of the Brain University of Oxford, Oxford, U. K. C35 Email: devlin@fmrib.ox.ac.uk Predictions Imaging results: Reading Words Discussion Background The current results are inconsistent with the hypothesis that the left occipito-temporal cortex is performing morphological decomposition at an orthographic level (3). The fact that non-affixed (spinach), apparently affixed (ponder) and affixed words (teacher), all show neural adaptation when paired with their embedded stems, suggests that this region processes abstract, visual form rather than apparent morphological structure. Together with previous findings (7, 9), we argue against the notion of either pre-lexical (10), lexical (8), or morphological (3) visual word forms stored in a particular patch of cortex. Instead, this region appears to integrate abstract, visual form with higher order properties of the stimulus such as meaning or sound (but not limited to these), based on functional (11, 12) and anatomical (13) connections to higher order association cortices (14). This account leads away from cognitive-based parcelations of cortex and towards an understanding of brain function in terms of information processing grounded in known anatomical and neurophysiological properties of the region. PMv The two hypotheses generate different predictions regarding activation of the left posterior fusiform region in visual masked priming: Morphological structure hypothesis: If orthographic representations are organised morphologically and stored in the left posterior fusiform gyrus, then one should observe neural priming (i.e. reduced activation) for pseudo-affixed (ponder-pond) and morphologically related word-pairs (teacher-teach) because both have apparent morphological structure. Orthographically related pairs such as spinach-spin should not show neural priming because they do not have apparent morphological structure. Visual form hypothesis: If the left posterior fusiform computes abstract visual form information more generally, then all three conditions (orthographically, pseudo-affixed, and morphologically related) pairs will show a neural priming effect relative to unrelated pairs as all shared equivalent orthographic overlap. Previously we have demonstrated that neural priming effects for morphologically related word pairs (e.g. teacher-teach) overlap with the adaptation observed for visually related words (e.g. ponder-pond) and semantically related words (e.g. idea-notion) (1). We argued that these results were consistent with the claim that morphology is an emergent property of interactions between form and meaning rather than a separate level of linguistic representation (2). Davis (3) noted, however, that the visual priming effect in the left posterior occipito-temporal cortex could be due to apparent morphological structure as the visually related word pairs looked as if they ended in a valid morpheme (e.g. ponder ≠ pond + -er). In other words, if morphemes are stored at a purely orthographic level (4), then the visual priming effect may have been due to repeated access to visual morphemes (3) rather than shared visual form more generally (1). pSTG PTr pOT aSTG Figure 5: Group results for unrelated words relative to consonant letter strings. Based on a mixed-effect group analysis shown on a single subject’s T1 image. • Activation was present in • the triangular (PTr) and opercular (POp) regions of Broca’s area, • ventral premotor cortex (PMv), • areas of the anterior and posterior superior temporal gyrus (a/pSTG), and • the posterior occipito-temporal region (pOT). Behavioural pre-testing • To verify the efficacy of the visual masking, a behavioural pre-test was conducted where participants were asked to read masked words aloud as best possible. • Words were presented for either 33 or 200msec and forward and backward masked with visual noise • Performance was at ceiling in for words presented for 200msec. In contrast, words presented for 33msec were only occasionally read aloud. • These results suggest that visually masked primes were not consciously perceived, consistent with post-hoc self-report in all experiments. Current Study Word reading in individuals • References • Devlin, J. T., Jamison, H. L., Matthews, P. M., & Gonnerman, L. M. (2004). Morphology and the internal structure of words. Proc Natl Acad Sci U S A, 101(41), 14984-14988. • Seidenberg, M. S., & Gonnerman, L. M. (2000). Explaining derivational morphology as the convergence of codes. Trends Cogn Sci, 4(9), 353-361. • Davis, M. H. (2004). Units of representation in visual word recognition. Proc Natl Acad Sci U S A, 101(41), 14687-14688. • Rastle, K., Davis, M. H., & New, B. (2004). The broth in my brother's brothel: morpho-orthographic segmentation in visual word recognition. Psychon Bull Rev, 11(6), 1090-1098. • Devlin, H., Devlin, J. T., Woolrich, M., Miller. K. and Jezzard, P. (2006) An efficient method for obtaining subject-specific HRF estimates in event-related fMRI. Poster at ISMRM 2006. • Cohen, L., Lehericy, S., Chochon, F., Lemer, C., Rivard, S., & Dehaene, S. (2002). Language-specific tuning of visual cortex? Functional properties of the visual word form area. Brain, 125, 1054-1069. • Devlin, J. T., Jamison, H. L., Gonnerman, L. M., & Matthews, P. M. (2006). The role of the posterior fusiform in reading. Journal of Cognitive Neuroscience, 18(6). • Kronbichler, M., Hutzler, F., Wimmer, H., Mair, A., Staffen, W., & Ladurner, G. (2004). The visual word form area and the frequency with which words are encountered: evidence from a parametric fMRI study. Neuroimage, 21(3), 946-953. • Price, C. J., & Devlin, J. T. (2003). The myth of the visual word form area. Neuroimage, 19(3), 473-481. • Dehaene, S., Cohen, L., Sigman, M., & Vinckier, F. (2005). The neural code for written words: a proposal. Trends Cogn Sci, 9(7), 335-341. • Bokde, A. L., Tagamets, M. A., Friedman, R. B., & Horwitz, B. (2001). Functional interactions of the inferior frontal cortex during the processing of words and word-like stimuli. Neuron, 30(2), 609-617. • Mechelli, A., Crinion, J. T., Long, S., Friston, K. J., Lambon Ralph, M. A., Patterson, K., et al. (2005). Dissociating reading processes on the basis of neuronal interactions. J Cogn Neurosci, 17(11), 1753-1765. • Distler, C., Boussaoud, D., Desimone, R., & Ungerleider, L. (1993). Cortical connections of inferior temporal area TEO in macaque monkeys. Journal of Comparative Neurology, 334(1), 125-150. • Price, C. J., & Friston, K. J. (2005). Functional ontologies for cognition: The systematic definition of structure and function. Cognitive Neuropsychology, 22(3-4), 262-275. • The aim of the current study was to evaluate these two hypotheses of left occipito-temporal involvement in orthographic processing • A visual masked priming paradigm with a lexical decision task was used to engage the region and fMRI was used to measure neural adaptation Figure 1: A schematic diagram of the visual masked priming paradigm. The prime was forward masked by a visual noise pattern and backward masked by the target so that participants were not consciously aware of the prime. • The design included four test conditions, two non-word conditions and a fixation baseline • Unrelated, orthographically related and pseudo-affixed pairs were matched for rated semantic similarity across conditions (1.0 = unrelated to 9.0 = identical meanings) • All related prime-target pairs were phonologically transparent in British English and had statistically equivalent orthographic overlap (mean = 4.4 letters, F2, 87<1) • Lexical stimuli were also matched for familiarity, frequency, concreteness, imageability and syllable length, while all stimuli were matched for letter length • Trials were presented in a pseudo-randomized, event-related design with “null events” and a mean ISI of approximately 5s • Sixteen native British English speakers (8F, 8M) participated in two runs with the order counter-balanced across subjects (B0=3T, gradient echo EPI, TR = 3sec, TE = 30msec, FOV = 192 x 256mm, matrix = 64 x 64) • Median RTs for correct responses were calculated per condition per subject and used in the behavioural analyses. Data from two subjects were excluded due to atypical performance: one had RTs > 200msec longer than any other participant and one fell asleep during the task • The functional imaging data were realigned to correct for small head movements, spatially smoothed with a 6 × 6 × 6mm FWHM Gaussian filter, and registered into standard space with an affine transformation • First level analyses used individually-tailored HRF estimates convolved with stimulus onsets to maximize sensitivity to BOLD signal change (5) • Briefly, run A was used to determine HRF shape and then used to analyse run B • Run B was used to determine the HRF shape for the analysis of run A Produced an unbiased, efficient estimate of individual subjects’ HRF • In each subject, a functional ROI was defined as a sphere (6mm radius) surrounding the peak activation for reading [Unrelated > fixation] in the left occipito-temporal area (a standard space sphere around [-42 -56 -18] ). The ROI was then used to evaluate priming effects A. Reading aloud paradigm B. Reading accuracy 98% #$%%#$ 500ms Accuracy (% correct) 1500ms reptile 33 or 200ms PASS $%##$% 200ms 500ms Time passive * 1000ms 33ms 9% $%##&@#@ 500ms Time * 1000ms 200ms 33ms Duration Figure 2: A. Schematic of the reading paradigm with visual noise patterns before and after the word. For short (33msec) but not long (200msec) durations, the masking blocked conscious awareness of the word. B. Results of the behavioural pre-test. Figure 6: In all cases, unrelated words relative to consonant strings activated left posterior occipito-temporal region near the occipito-temporal sulcus (white arrows). A dotted line marks the collateral sulcus between the fusiform and parahippocampal gyri. Table: Sample stimuli in the lexical decision task Semantic Condition Example Similarity Rating 1. Unrelated journal – HAZE 1.3 2. Orthographic spinach – SPIN 1.4 3. Pseudo-affixed planet – PLAN 1.5 4. Morphological poetry – POET 7.6 5. Pseudowords florze – HALDA – 6. Consonant letter strings pchmmv – WLPBX – Demonstrates that the lexical decision task engages the specific occipito-temporal region observed in previous studies of the “visual word form area” (6-9) Behavioural results: Lexical decision • Accuracy across conditions was 97.3%, indicating that subjects had no difficulty performing the task. • There was a significant main effect of Relation (Unrelated, Orthographical, Pseudo-affixed, Morphological) between prime and target on reaction times (F3,39=9.1, p<0.001). • Post-hoc comparisons revealed that the only significant priming effect was for morphologically related words (16 msec priming, t13=2.7, p=0.05 after Bonferroni correction for multiple comparisons, two-tailed). Neural priming effects Words Nonwords 99% 99% 98% 97% 97% • For each individual, the mean percent BOLD signal change relative to fixation was computed per condition: 93% Accuracy (% correct) Words Nonwords * * Email: devlin@fmrib.ox.ac.uk * 0.63 Unrel Orth PsAff Morph PsWord Cons 0.56 BOLD signal change (%) 0.41 Figure 3: Accuracy to words (left) and nonwords (right) in the lexical decision task. There were no significant differences between lexical conditions in accuracy (F3,39=2.0, p=0.13) 0.37 0.36 0.36 Unrel Orth PsAff Morph PsWord Cons Data Analyses Words Nonwords Figure 7: Neural repetition priming effects. There was a significant difference between lexical conditions in BOLD signal change (F3,39=3.9, p=0.015) +25ms +21ms * –16ms • Within the left occipito-temporal region activated by words, all three conditions which shared orthographic overlap showed a significant neural priming effect: • Orthographic (spinach-SPIN) t13=3.2, p=0.011 • Pseudo-affixed (planet-PLAN) t13=3.0, p=0.017 • Morphological (poetry-POET) t13=2.7, p=0.027 • The effects were case-independent, consistent with previous studies (6, 7). • In other words, words which shared visual form, independent of apparent morphological status, led to a significant reduction in BOLD signal relative to unrelated words. Reaction Times (msec) Unrel Orth PsAff Morph PsWord Cons Figure 4: RTs for words (left) and nonwords (right). The magnitude of the priming effect relative to unrelated pairs is indicated above each condition in msec. The * indicates a significant priming effect at p=0.05.






















