Download

1 / 36
430 likes | 1.23k Views
Alele multiple (polialelia ). Alelele pot fi definite şi ca alternative ale aceleia ş i gene, control â nd expresii diferite ale unui anumit caracter al organismului. In urma muta ţ iei unei gene de tip s ă lbatic care de regul ă este dominant ă (A) apare alela sa recesiv ă (a).

E N D
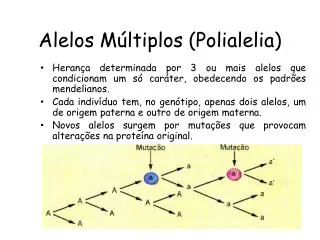
Alele multiple (polialelia) • Alelele pot fi definite şi ca alternative ale aceleiaşi gene, controlând expresii diferite ale unui anumit caracter al organismului. • In urma mutaţiei unei gene de tip sălbatic care de regulă este dominantă (A) apare alela sa recesivă (a). • Mutaţia este mult mai rarăîn sens invers (a A). • Procesul mutaţional s-a desfăşurat progresiv în timp şi a dus uneori la apariţia succesivă a mai multor alele (a1, a2, a3,....an) care controlează expresia aceluiaşi caracter. • Asemenea gene poartă denumirea depolialele sau alele multiple.
Organismele diploide prezintă prin definiţie numai două alele la un moment dat. • Acest aspect este datorat prezenţei cromozomilor omologi sub formă pereche, fiecare din cei doi cromozomi omologi conţinând doar una din cele două alele ale unei perechi de gene. • Fenomenul polialeliei este un fenomen populaţional, ceea ce înseamnă că la unii indivizi ai populaţiei, pe o pereche de cromozomi se află alelele AA: la alţi indivizi ai populaţiei pe aceeaşi pereche de cromozomi omologi se va afla perechea a1a1, la alţii a2a2, la alţii a3a3, s.a.m.d.
Un sistem de alele multiple este întâlnit la omîn cazul determinării grupelor de sânge în sistemul ABO. • Apariţia grupelor diferite de sânge în acest sistem este datorată existenţei a trei alele ale aceleiaşi gene (LA, LBşi l) care pot determina apariţia a patru grupe sangvine diferite: • A (LALA sau LAl); • B (LBLB sau LBl), • AB (LALB) şi • O (ll). • Apariţia acestor grupe sangvine diferite este datorată prezenţei a doi antigeni diferiţi (A şiB)
Relaţiile de dominanţă între alelele sistemului ABO pot fi reprezentate astfel: LA > l, LB > l, LA = LB; Alelele LAşi LBsunt ambele dominante asupra lui l iar împreună sunt codominante. • Fenotipul AB este datorat prezenţei ambelor alele dominante (LA şi LB), indivizii prezentând pe suprafaţa hematiilor ambii antigeni (A şi B). • Un individ cu genotipul ll, nu va avea pe suprafaţa hematiilor niciunul din antigenii A şi B şi va avea grupa O. • Modul de moştenire a caracterelor determinate de alele multiple nu diferă de cel al caracterelor determinate de două alele.
Un exemplu de alele multiple întâlnim la D.melanogasterîn cazul genelor care determină culoarea ochilor. • Pentru această genă una din primele mutante cunoscute a fost numităwhite (w) deoarece ducea la apariţia ochilor de culoare albă ca urmare a lipsei pigmenţilor din ochi. • Ulterior au fost identificate numeroase alte alele care aveau un efect fenotipic intermediar între culoarea roşie normalăşi cea albă. • Aceste aspecte fenotipice variate şi caracteristice pentru fiecare alelăîn parte erau datorate unei cantităţi diferite de pigmenţi oculari, cantitate caracteristicăînsă pentru fiecare mutantăîn parte.
Interacţiunea genelor • In cazul experimentelor de dihibridare efectuate de G.Mendel perchile de alele se comportau independent unele de altele. • Independenţa privea două aspecte: • Separarea în gameţi a celor două alele ale unei perechi se realiza independent de separarea alelelor celeilalte perechi; • Cele două perechi de alele erau independente în ceea ce priveşte expresia fenotipică.
Adesea perechile de alele se separă independent unele de altele, în gameţi dar nu acţionează independent în ce priveşte expresia fenotipică. • Altfel spus, efectul fenotipic al unor gene de la un locus depinde de prezenţa unor gene din alţi loci. • Acest tip de interacţiune între efectele unor gene din loci diferiţi (gene nealele) este denumită interacţiune genică. • În cazul interacţiunii genice, produşii unor gene din loci diferiţi concură în producerea unor fenotipuri noi care nu pot fi explicate prin participarea unei singureperechi de alele
Un exemplu interesant de interacţiune a genelor care nu determină modificarea raportului mendelian de segregare, ci numai apariţia de fenotipuri noi, deosebite de ale genitorilor, a fost observat la încrucişarea între rase de găini cu tipuri diferite creastă. Trandafir mazăre simpla nucă
Acest caracter s-a dovedit a fi determinat de două gene nealele R şi P. • Există patru tipuri alternative de creste : simplă(rrpp), mazăre (rrPP), trandafir (RRpp), şi nucă(RRPP). • Când se fac încrucişări trandafir x simplă sau mazăre x simplă se obţin următoarele rezultate :
Aceste rezultate sunt tipice pentru segregarea de tip Pisum cu respectarea deci a legităţilor mendeliene (trandafir şi mazăre fiind ambele dominante asupra genei care determină forma simplă a crestei, formă care apare datorită stării de homozigoţie recesivă). • In urma încrucişării trandafir x mazăre însă, în F1 toţi indivizii vor avea creasta tip nucă. In F2 vor apărea toate tipurile de creastăîn următoarele proporţii : 9/16 nucă, 3/16 trandafir, 3/16 mazăre şi 1/16 simplă. • Din aceste încrucişări reiese faptul că din interacţiunea celor două gene dominante R şi P apare un nou tip de creastă, deosebit de cel al genitorilor, şi anume tipul nucă.
Când două sau mai multe genenealele acţionează asupra aceluiaşi caracter, o genă poate masca efectul celeilalte gene, cam în aceeaşi măsurăîn care efectul unei gene recesive este mascat de alela dominantă. • Acest fenomen prin care o genă poate masca sau suprima efectul unei alte gene nealele cu ea, a fost denumit epistazie. • Gena epistatică este o genă care aparţine unei perechi de alele şi împiedecă manifestarea alelei dominante dintr-o altă pereche de alele. • Gena supresată poartă numele de genăhipostatică.
Fenomenul de epistazie este implicat în aproape toate tipurile de interacţiune genică. • Dominanţa implică supresia genică intra-alelică (mascarea expresiei unei alele recesive de către alela dominantă). • Epistazia cuprinde supresia genică inter-alelică (mascare expresiei unei gene se către o altă genă nealelă cu ea). • Raportul fenotipic clasic 9:3:3:1 observat la descendenţii unor părinţi dihibrizi se modifică datorită epistaziei în raporturi care reprezintă combinaţii ale acesteia. • Gena epistatică poate fi dominantă sau recesivă.
Gene epistatice dominante (12:3:1) • Fenomenul epistaziei dominante a fost observat la o încrucişare între două varietăţi de Avena sativa, una cu boabe negre şi alta cu boabe galbene.
Cele 12 combinaţii care conţin gena A în stare homozigotă sau heterozigotă sunt de culoare neagră; • 3 combinaţii care au gena dominantă B în stare homozigotă sau heterozigotă, dar nu conţin gena A sunt de culoare cenuşie, • Combinaţia cu ambele gene recesive (ab) produce boabe galbene. • Din cele de mai sus rezultă faptul că gena dominantăA împiedecă expresia genei dominante Bşi este deci epistatică pe când gena dominantă,a cărei acţiune este inhibată este hipostatică.
Gene epistatice recesive(9:4:3) • Un exemplu de epistazie recesivă ni-l oferă ereditatea culorii blănii la anumite rase de câini. La încrucişarea între două rase de câini, una homozigotă albă cu una homozigotă maron se obţin în F1 numai descendenţi de culoare neagră. Incrucişările indivizilor din F1 au produs descendenţi în raportul de segregare de : 9 negru : 4 galben : 3 maron.
În acest caz alelele B şi b produc culoarea neagră şi respectiv maron, dar numai în prezenţa alelei dominante C. • Pentru a distinge această interacţiune de o relaţie dominanţă şi recesivitate, se spune căcc este epistatic faţă de B sau b; adică homozigoţia alelei c previne formarea oricărui pigment, indiferent de alelele pentru alte culori. • Efectul pe care alelele cc îl au asupra lui B poartă numele de epistazie recesivă.
In acest caz c este o genă epistatică recesivă deoarece este nevoie de două copii ale acestei gene pentru a masca pigmenţii maron şi negri • Acest caz de epistazie nu este cauzat de o blocare a căii de sinteză a pigmentului negru • Câinii galbeni pot produce pigmentul maron sau negru aşa cum se poate vedea la nivelul nasului şi buzelor • Gena c acţionează prin blocarea depunerii pigmentului în blană
Mecanismul molecular al epistaziei recesive la Collinsia parviflora
Gene duplicate în epistazia recesiva • Albinismul se caracterizează prin lipsa pigmentului şi este o caracteristică genetică întâlnită la multe plante şi animale. • Producerea pigmentilor se face aproape întotdeauna printr+un proces biochimic în mai mulţi paşi; albinismul poate deci implica un fenomen de interactiune a genelor. • La melcul de apă dulce Physa heterostroha, albinismul poate rezulta ca urmare a prezenţei a oricăror alele recesive de la doi loci diferiţi. • Când doi melci albino se încrucişează, toată generaţia F1 este pigmentată.
În F2 9/16 din descendenţi sunt pigmentaşi iar 7/16 sunt albino. • Raportul de 9:7 este generat probabil de producerea pigmentului într-un proces în doi paşi. • Pigmentul (compusul C) este produs numai după ce compusul A a fost transformat în produsul B de enzima I şi după ce produsul B a fost transformat de enzima II în produsul C. • Raportul de 9:7 apare când alele dominante la ambii loci (A_B_) produc pigment; orice alt genotip duce la apariţia melcilor albino. • Albinismul este datorat lipsei compusului C. • În acest caz a este epistatică faţă de B şi b faţă de A; ambele sunt gene epistatice recesive deoarece e nevoie de prezenţa ambelor copii ale oricărei din cele doua alele pentru a bloca producerea de pigment.
Epistazia constă în mascarea expresiei unei gene de către o altă genă de la un locus diferit de al primei (genă nealelă). • Gena epistatică este cea care maschează expresia unei gene nealele. • Gena hipostatică este cea a cărei aspect este mascat. • Genele epistatice pot fi dominante sau recesive.
În fiecare exemplu prezentat s-a observat o modificare a raportului de segregare de 9:3:3:1. • Modul de moştenire a genelor ce produc aceste caracteristici nu diferă de cel al genelor ce pentru caractere genetice simple. • Principiile mendeliene ale segregării independente se aplică şi în aceste cazuri. • Diferenţa constă în modul în care produşii genelor interacţionează pentru a produce fenotipul. • Proporţiile fenotipice sunt exprimate în şaisprezecimi deoarece sunt implicate două perechi de alele ce segregă independent. • În general proporţia descendenţilor este de (1/2)2n, unde n reprezintă numărul de loci ce segregă independent
Complementarea: determinarea faptului că o mutaţie apare la un locus sau la doi loci diferiţi • La Drosophila melanogaster white este o mutaţie x-linkată care produce ochi de culoare albă. • Apricot este tot o mutaţie x-linkată care duce la apariţia ochilor de culoarea caisei (portocaliu deschis). • Mutaţiile white şi apricot au apărut în acelaşi locus sau în loci diferiţi? • Putem utiliza testul de complementare pentru a răspunde la această întrebare.
Testul este uşor de aplicat. Se încrucişează două forme mutante şi se analizează F1. • Dacă în F1 apare fenotipul sălbatic putem concluziona că mutaţiile au loc în gene alele diferite care îmreună contribuie la realizarea fenotipului. • Explicaţie: dacă două gene nealele sunt implicate fiecare genă va avea o mutaţie în una din alele în timp ce cealaltă va fi sălbatică (nemutantă). Deoarece în F1 se vor exprima ambele alele necesare formei sălbatice fenotipul va fi corespunzător formei sălbatice
Dacă în F1 nu apare fenotipul sălbatic ci unul mutant, putem concluziona că mutaţiile au loc în interiorul aceleiaşi gene. • Explicaţie: • dacă mutaţiile există în poziţii diferite ale aceleiaşi gene, în F1 fiecare cromozom omolog va prezenta câte o variantă a mutaţiei; • In lipsa produsului genei sălbatice va apărea un fenotip mutant.
Moştenirea caracterelor continue • Pâna acum am trecut în revistă carectere care manifestau doar câteva fenotipuri distincte. • În experimentele lui G.Mendel seminţele de mazăre erau: netede sau zbârcite, galbene sau verzi. • În sistemul ABO există 4 grupe de sânge distincte: A, B, AB, O • Astfel de caractere care determină doar câteva fenotipuri distincte au fost numite caractere discontinui.
Nu toate caracterele prezintă fenotipuri discontinue. • La om, greutatea corporală şi talia (înălţimea) sunt astfel de caractere deoarece nu întâlnim greutăţi sau înălţimi distincte ci un şir gradat de fenotipuri de la cele mai mici valori până la cele mai mari. • Astfel de caractere care prezintă o distribuţie continuă a fenotipurilor au fost denumite caractere continue • Deoarece aceste caractere prezintă multe fenotipuri şi trebuie descrise în termeni cantitativi au fost denumite şi caractere cantitative.
Caracterele continui apar, cel mai adesea, datorită faptului că gene din mai mulţi loci diferiţi interacţionează pentru a produce un anume fenotip. • Când un caracter este codificat de două alele de la un singur locus, există doar 3 genotipuri posibile. • Când sunt implicaţi doi loci cu două alele fiecare, vor fi 32=9 genotipuri posibile. • Când caracterul este determinat de 8 loci, fiecare cu două alele, sunt posibile 38 = 6561 genotipuri diferitepentru acel caracter. • Numărul de genotipuri posibile pentru un caracter este egal cu 3n unde n este egal cu numărul de loci cu două alele care contribuie la determinarea caracterului respectiv.
Caracterele acăror determinara este datorată mai multor perechi de alele din loci diferiţi şi interacţiunii lor cu mediu, au fost denumite caractere poligenice multifactoriale. • La om, numeroase caractere normale (inteligenţă, talie, greutate, culoarea pielii, a ochilor) sau patologice (autismul, cancerul, diabetul, hipertensiunea, schizofrenia etc.) au o astfel de determinare.