Download
1 / 28
280 likes | 296 Views
This text discusses the mechanisms of maternal recognition of pregnancy and the maintenance of the corpus luteum (CL) in mares. It explores the role of prostaglandin F2a (PGF2a) in CL regression and the importance of the critical period for maternal recognition of pregnancy. The text also compares the utero-ovarian pathway in mares to that in cows and ewes.

E N D
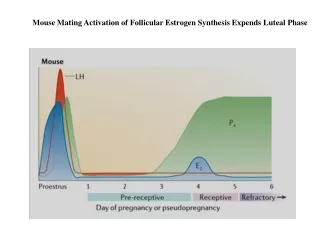
Mouse Mating Activation of Follicular Estrogen Synthesis Expends Luteal Phase
Maternal Recognition of Pregnancy in the Mare In the cyclic mare the CL starts regression on about 14 days post-ovulation. CL regression and declining plasma progesterone concentrations are associated with increased plasma and uterine PGF2a secretion. Stabenfeldt et al. In:Prostaglandins in Animal Reproduction, Acta Vet Scad. 1981, Suppl 77:159
Hysterectomy prior to Day 12 to 14 prolongs CL function for 70 to 140 days. Removal of the equine conceptus by flushing the uterine horns prior to Day 15 results in luteolysis, but after Day 15 the CL function is maintained. Therefore, Day 14 to 16 is the critical period for maternal recognition of pregnancy. Conceptus removal on Day 10, 15 and 20 resulted in interestrous intervals of 22, 38 and 46 days respectively, compare to 24 days in cyclic controls. Possible Mechanism for Maintenance of CL 1. Prevent PGF2a synthesis and release. 2. Prevent PGF2a from entering the maternal vasculature. 3. Prevent PGF2a from binding to luteal cell membrane receptors. 4. Stimulate luteal gonadotropin receptors.
The pathway of PGF2a in reaching the ovaries in the mare differs from cow and ewe. There is no direct unilateral effect of the luteolysin from the uterus to the ovaries. The uterine artery is relatively straight and caudal to the ovarian vein, as it passes obliquely across the ovarian vein. In the mare, CL regression appears to occur through a systemic pathway. PGF2a in the utero-ovarian venous plasma increases on Day 14 in cyclic mares. However, PGF2a is much lower in pregnant mares. PGF2a Binding by CL Day 14 luteal PGF2a receptors reach the highest level. PGF2a binding rapidly declines after Day 14 in cyclic mares but continues to increase up to Day 18 in pregnant mares.
There is a significant reductionPGF2a luminal content in pregnant compared to cyclic mares. Evidence indicates that in the presence of the conceptus, PGF2arelease or synthesis is inhibited Collected by cervical lavage Endometrial Production ofPGF2a Nonpregnant mares, PGF2a endometrial content and synthesis increases on Day 14-16. Pregnant mares synthesis continues to increase to Day 20, however content is lower until Day 18-20. Reproductive Biology of the Mare, Ed. O.J. Ginther, pp419-456
With systemic route of PGF2a in the mare the conceptus must migrate between the horns to inhibit oxytocin receptor and the release and/or synthesis of PGF2a.
Twining Tree Ginther, IN: Reproductive Biology of the Mare, 2nd Ed. 1992
How is Embryo Reduction Accomplished? Ginther, IN: Reproductive Biology of the Mare, 2nd Ed. 1992
Maternal Recognition in the Ewe Uteroovarian Pathway In comparison to the gilt and mare, regulation of the uterus on CL function in the ewe and cow is exerted through a local utero-ovarian pathway. Experiments involving surgical anastomoses of a main uterine vein or ovarian artery to the corresponding vessel on the opposite side have demonstrated that the local utero-ovarian pathway is the mechanism of CL regression in the cow and ewe. Unilateral Hysterectomy: When the uterine horn is ipsilateral to CL, regression occurs more readily. Unilateral Pregnancy: Establishment of pregnancy in one side of the uterus by surgical isolation of the uterine horns results in luteal maintenance on the gravid side more readily than on the nongravid side in sheep and cattle.
A. Removal of the uterine horn maintains the CL in contrast to a intact uterine vein. Demonstrates utero-ovarian pathway. B. CL will regress if the uterine vein of the intact uterine horn is anastomosed with the vein on the contralateral side. Note the fusion is above the ovarian artery! C. Ovarian artery from the uterine intact side anastomosed with ovary from the side lacking uterine horn. Note anastomosed artery is cut at the region above were the artery touches the uterine vein. O.J. Ginther, In:Prostaglandins in Animal Reproduction, Acta Vet Scad. 1981, Suppl 77:103
A. The presence of the conceptus prevents CL regression on the gravid side with loss of CL on the surgically isolated nongravid horn. B. The CL on the nongravid side is rescued if the uterine vein from the gravid horn is anastomosed to the vein on the nongravid horn. C. The CL on the nongravid side is rescued if the ovarian artery from the gravid horn is anastomosed to the ovarian artery on the nongravid horn. Note: position of anastomoses. O.J. Ginther, In:Prostaglandins in Animal Reproduction, Acta Vet Scad. 1981, Suppl 77:103
When the ovarian artery from the nongravid side was anastomosed with the ovary of the gravid horn the CL regressed. Indicating the conceptus factor to maintain the CL is not systemic. When the ovarian artery of the nongravid side is anastomosed at a position where the conceptus factor can be transferred with it to the ovary, the CL is maintained. Suggests a factor that can have a protective role to PGF2a which is a function of PGE. O.J. Ginther, In:Prostaglandins in Animal Reproduction, Acta Vet Scad. 1981, Suppl 77:103
Maintenance of CL Function in Ewe and Cow Although luteolysis occurs through PGF2a action, the effect of the embryo on PGF2a secretion in the uterus in early pregnancy is conflicting. Plasma levels of PGF2a have been reported to be diminished, unchanged or higher during early pregnancy compared to the cyclic ewe. Recent evidence by McCracken indicates rising spikes of PGF2a at Day 13 and 14 in both pregnant and nonpregnant ewes. However, major luteolytic pulses seen on Days 14, 15 and 16 are absent in pregnant ewes. Nonpregnant 5 pulses within 24 hours It takes five hour-long infusions of PGF2a (0.4 to 1.2 mg/h) over 25 hours to cause complete CL regression in the ewe. Pregnant McCracken et al. 1981, Acta vet scand, Suppl 77:71
Myometrium Day 0 Day 0 Progesterone receptor in the endometrium Note presence of PR in epithelium, stroma and myometrium on Day 5. Day 15 Note absence of PR in epithelium on Day 15 of the estrous cycle or pregnancy
McCracken Model of Oxytocin Control of CL Regression McCracken et al. 1981, Acta vet scand, Suppl 77:71 1. Because of declining progesterone action on the uterus towards the end of the luteal phase, estradiol-17b is now able to stimulate formation of endometrial oxytocin receptors. (Takes 6 hours) 2. Endogenous levels of oxytocin (posterior pituitary) interact with the receptor. 3. PGF2a secretion from the uterus follows oxytocin stimulation. 4. Progesterone secretion declines with luteolysis as result of secretion, locally transferred to the ipsilateral ovary. 5. More oxytocin is released with PGF2a stimulation of the CL to reinforce more uterine PGF2a release. 6. Since oxytocin receptors will down-regulate, subsequent hour-long episodes of PGF2a release at intervals of 6 h will allow replenishment of oxytocin receptor.
1. Loss of P action occurs simultaneously in the hypothalamus and endometrium thus returning E action. 2. E from follicles can then stimulate increase pulses of OT from hypothalmus with increase in endometrial OTR. 3. Subluteolytic pulses of PGF2a will be releasedfrom uterus following stimulation by post pit OT with uteine OTR. 4. Subluteolytic levels of PGF2a will bind high affinity PGF2a receptors (HFRR) on large luteal cells to release OT. 5. Release of luteal OT will cause a luteolytic release of PGF2a from the endometrium. 6. The high release of PGF2a now binds to low and high affinity receptors to cause inhibition of progesterone secretion and release of more OT. Positive feedback loop. 7. Next release of luteolytic PGF2a depend on release of post pit OT, recovery of receptors for PGF2a after 6-9 hours and recovery of uterine OTR. McCracken Model of Oxytocin Control of CL Regression McCracken et al. 1981, Acta vet scand, Suppl 77:71 Custer et al. Reprod Dom Anim 31:449
Cloned gene sequence of oTP-1 and bTP-1 are homologous to bovine a II interferon family. Interferons are antiviral, antiproliferative and immunomodulatory The protein is now classified as ovine or bovine interferon-tau (oIFNt) IFNt increases the uterine expression of: 1) 2’, 5’ oligoadenylatesynthetase 2) b2-microgolubin 3) Interferon regulatory factor-1 and -2 4) Ubiquitin cross-reactive protein 5) Mx protein 6) Graulocytechemotactic protein-2 Binds to the IFN type I receptors on the surface and glandular epithelium of the endometrium. It clear that IFNt is involved with maternal recognition of pregnancy in ruminants. Binds to cytosolic proteins to prevent proteasomal degradation (prevent apoptosis) Induced by type I IFN, antiviral protein - Possibly can act as GTPase
Schematic of current working hypothesis on cell-specific IFNT signaling in the endometrium of the ovine uterus. IFNT, produced by developing conceptuses of ruminants, binds to IFNAR present on cells of the ovine endometrium. (A) In the uterine luminal and superficial glandular epithelia (LE/sGE), IRF2, a potent and stable transcriptional repressor, increases during early pregnancy. The continual presence of IRF2 inhibits classical IFN-stimulated genes or ISGs (STAT1, STAT2, IRF9, B2M, ISG15, MHC, and OAS) through direct ISRE and IRFE binding and coactivator repulsion. Thus, critical factors in the classical JAK-STAT-IRF pathway (STAT1, STAT2, and IRF9) are not present, resulting in the absence of ISGF3 or IRF1 transcription factors necessary to transactivate ISGs. However, IFNT does activate an unknown cell signaling pathway that results in induction of WNT7A and stimulation of non-classical IFNT-stimulated genes (CST3, CTSL, and LGALS15) specifically in LE/sGE. (B) In cells of the stroma (ST) and middle to deep GE, the IRF2 repressor is not expressed. Thus, IFNT-mediated association of IFNAR subunits facilitates cross-phosphorylation and activation of JAK1 and Tyk2, which in turn phosphorylates the receptor and creates a docking site for STAT2. STAT2 is then phosphorylated, thus creating a docking site for STAT1 which is then phosphorylated. STAT1 and STAT2 are then released from the receptor and can form two transcription factor complexes, GAF and ISGF3. ISGF3 is formed by association of a STAT1-2 heterodimer and IRF9 in the cytoplasm, translocates to the nucleus, and transactivates genes containing an ISRE(s), such as STAT1, STAT2, IRF9, B2M, ISG15, MIC, and OAS. GAF is formed by STAT1 homodimers, which translocates to the nucleus and transactivates genes containing a GAS element(s) such as IRF1. IRF1 can also bind and transactivate ISRE-containing genes as well as IRFE-containing genes. The simultaneous induction of STAT2 and IRF9 by IFNT appears to shift transcription factor formation from GAF towards predominantly ISGF3. Therefore, IFNT activation of the JAK-STAT-IRF signal transduction pathway allows for constant formation of ISGF3 and GAF transcription factor complexes and hyperactivation of ISG expression in the ST and GE. B2M, β-2-microglobulin; CST3, cystatin C; CTSL, cathepsin L; GAF, -activated factor; GAS, activation sequence; IFNAR, type I IFN receptor; IFNT, interferon; IRF1, interferon regulatory factor 1; IRF2, interferon regulatory factor 2; IRFE, IRF-response element; IRF9, IFN regulatory factor 9 (alias ISGF3G); ISG15, (ISG15 ubiquitin-like modifier; alias IFI15 or UCRP); ISRE, IFN-stimulated response element; JAK, janus kinase; LGALS15, galectin 15; MIC, MHC class I polypeptide-related sequence; OAS, 2',5'-oligoadenylate synthetases; STAT1, signal transducer and activator of transcription 1, 91 kDa; STAT2, s ignal transducer and activator of transcription 2, 113 kDa; WNT7A, wingless-type MMTV integration site family, member 7A. Spencer et al. 2008; Reproduction135: 165 - 179.