Download

1 / 53
741 likes | 1.61k Views
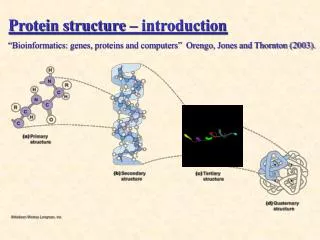
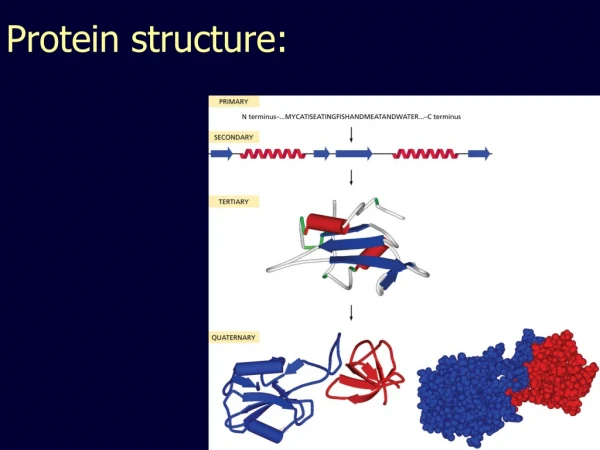
Introduction to Protein Structure. Q: Whats that?. A: Something, you get Noble prize for. John Kendrew & Max Perutz 1962 Structures of myoglobin & hemoglobin. Subjects, covered in this lecture. Amino acids and their properties Peptide geometry Secondary structure Motifs Domains

E N D
Introduction to Protein Structure Q: Whats that? A: Something, you get Noble prize for... John Kendrew & Max Perutz 1962 Structures of myoglobin & hemoglobin
Subjects, covered in this lecture • Amino acids and their properties • Peptide geometry • Secondary structure • Motifs • Domains • Quaternary structure
Why bother about protein structure? • Gives you an visual image of how proteins look like. • Study of protein structures allows to gain an insight into how protein really accomplish their function. • Nobel prizes...
Amino Acids • 20 different ones, sharing a common backbone but varying side chain. • Classed according to their chemical properties • L-form
nonpolar amino acids • R group consists of carbon chains leucine and isoleucine are structural isomers
nonpolar amino acids - R group consists of carbon chains Phenylalanine and tryptophan have aromatic rings which are flat due to the double bond network Tryptophan is often classified as being polar because of the NH group. In practice, however it has more of hydrophobic properties Proline has its R group bound to the amino nitrogen to form a ring network Methionine has a sulphur atom in its sidechain sulphur has the same valence as oxygen
polar amino acids - R group consists of carbon, oxygen and nitrogen atoms together they make the sidechain more hydrophilic Ser and thr are a mix of carbon chains and hydroxyl functional groups (-OH). Cysteine has a thiol group (-SH) which is otherwise structurally similar to serine but not chemically similar Asn and gln have an amide functional group
charged amino acids - R group has a charge at physiological pH (7.4). pK of the charged groups vary carboxyl group carboxyl group guanidinio group imidazole group, sometimes charged Most often classified as a polar amino acid amino group
Cysteine and disulphides • The almost exclusively only way to covalently link two non-sequential residues is by forming a disulphide bridge • Formation of disulphide requires an oxidative environment, threfore disulphides are very rare in intracellular proteins but quite abundant in secretory proteins
Peptide units • A peptide is a set of covalently bonded amino acids. • The covalent bond is usually referred as peptide bond
Structural biologist’s peptide unit – from Ca to next Ca - all main chain atoms within the unit lie in a plane Biochemist’s peptide unit –from N to C – all main chain atoms within the unit lie in the same residue
Angles phi (f) N-Ca • Angle psi (y) Ca-CO • Angle omega (w) C-N
The w angle, cis- and trans- peptides • Because of the partly double nature of peptide bond, w is always close to 180o for trans- peptides or 0o for cis- peptides (±30o in exterme cases) • Cis- peptides are energetically extremely unfavourable (~1000 fold) because of steric clashes between the neighbouring Ca atoms
The only exception is peptide bond before proline, where cis- peptide is just 4 times less favourable than trans- peptide, because there are some steric clashes in both cis- and trans- forms • Proline cis-trans isomerization is an important factor in protein folding, which is why there are special enzymes – prolylpeptydyl isomerases to catalyze the transition from one form to another
According to statistics, 0.03% of non-proline peptides and 5.2% of X-Pro peptides are in cis- conformation, resulting in a total of 0.3 % cis-peptides • In most cases cis- peptides, especially non-proline, occur for a good reason, for example to maintain some particular conformation in the active site of enzyme
Main chain conformations (I) • Only certain combinations of y and f are allowed, due to steric clashes of backbone atoms and Cb atom. Plot of these combinations yields the Ramachandran plot. • All amino acids clusters in specific regions (called allowed regions) except Gly (explains why Glycine is an important amino acid).
In good quality structures only about 2% of amino acid residues are found in the disallowed regions of Ramachandran plot • Of course, residues with disallowed conformations often have some important function in proteins
Side chain conformations • Side chains can have in principle different conformations (rotation of Ca-Cb...) • The observed conformations in protein structures are the ones which are more energetically favourable (rotamers).
Name three amino acids which are very different from others! Proline • No free amino group • Very rigid • Introduces breaks in a helices and b strands Glycine • Lacks a side chain • Can be found anywhere in Ramachandran plot • In proteins often found in flexible regions with unusual backbone conformations Cysteine • Disulphides
The hydrophobic core • The hydrophobic sidechains of protein has a tendency to cluster together in order to avoid unfavourable contacts with polar water molecules • As a result, in general, hydrophobic sidechains are located in the interior of protein, forming the hydrophobic core • Polar and charged amino acids usually are located on the surface of the protein • Polar and charged residues also can make hydrophobic contacts with their aliphatic carbon atoms • Polar and charged residues are seldom completely buried within the core and even when they are, the polar groups are almost invariably involved in hydrogen bond formation
The reasons of secondary structure formation • Since sidechains of hydrophobic residues are located in the hydrophobic core, the mainchain atoms of the same residues in most cases are also within the hydrophobic core • Since the presence of polar groups in hydrophobic environment is very unfavourable, the main chain N- and O- atoms have to be neutralised by formation of hydrogen bonds • The two most efficient ways of hydrogen bond formation is to build an alpha helix or a beta sheet
The alpha helix • 3.6 residues per turn • the hydrogen bonds are made between residues n and n+4
Variants of alpha helix • In regular a helix, residue n makes a H-bond with residue n+4 • In 310 helix, residue n makes a H-bond with residue n+3. There are 3 residues per turn, connected by 10 atoms, hence the name 310 • In p helix, residue n makes a H-bond with residue n+5 • In p helix there is a hole left in the middle of helix and in 310 helix the main chain atoms are packed very tightly. None of above is energetically favourable • 310 and especially p helices occur rarely and usually only at the ends of regular a helix or as a separate single-turn helix
Handedness of alpha helix • The a helices as well as 310 and p helices ale almost exclusively right-handed • In very rare occasions, left handed a and 310 helixes can occur. They are always very short (4- 6 residues) and normally involved in some important function of protein like in active site or ligand binding • There are about 30 reported cases of left-handed helices. In contrast, the number of known right handed helices is of order of hundreds of thousands
Good and bad helix formers • Different side chains have been found to have weak but definite preferences for helix forming ability • Ala, Glu, Leu and Met are good helix formers • Pro, Gly, Tyr and Ser are very poor helix formers • The above preferences are not strong enough to be used in accurate secondary structure predictions
Periodic patterns in a helices • The most common location of an a helice is along the outside of protein, with one side of the helix facing the hydrophobic core and other side facing the solvent • Such a location results in a periodic pattern of alterating hydrophobic and polar residues • On itself, however, the pattern is not reliable enough for structure prediction, since small hydrophobic residues can face the solvent and some helices are completely buried or completely exposed
Beta sheets Antiparallel Parallel
A mixed b sheet A mixed b sheet is far less common than antiparallel or parallel
Twist in b sheets • Almost all b sheets in the known protein structures are twisted • The twist is always right-handed
Loops • Loops connect secondary structure elements • Loops are located on the surface of protein • In general, main chain nitrogen and carbonyl oxygen atoms do not make H-bonds each to other in loops • Loops are rich in polar and charged residues • The lenght of loops can vary from 2 to more than 20 residues • Loops are very flexible, which makes them difficult to see in either x-ray or NMR studies of proteins • Loops frequently participate in forming of ligand binding sites and enzyme active sites • In homologous protein families loop regions are far less conserved than secondary stucture elements • Insertions and deletions in homologous protein families occur almost exclusively in loop regions
Hairpin loops and reverse turns • Loops, which connect two adjacent antiparallel beta strands are called hairpin loops • 2 residues long hairpin loops are often called reverse turns, beta turns or simply turns Type I turn Type II turn Hairpin loop Strand1 Strand2
Motifs • Simple combinations of a few secondary structure elements occur frequently in protein structures • These units are called supersecondary structure or motifs • Some motifs can be associated with a specific biological function (e.g. DNA binding) • Other motifs have no specific biological function alone, but are part of larger structural and functional assemblies
Helix-loop-helix motifs DNA binding motif Calcium binding motif
The hairpin b motif • Two adjacent anti-parallel b strands, joined by a loop • The hairpin motif can occur both as an isolated unit or as a part of bigger b sheet Snake venom- erabutoxin Bovine trypsin inhibitor
24 different ways to connect two b hairpins • Only the first 8 arrangements exist in known proteins
The Greek key motif • The most common way to connect 4 adjacent antiparallel b strands The Greek key motif in Staphilococcus nuclease
The b-a-b motif • A convinient way to connect two paralel beta strands • b-a-b motif is a part of almost all proteins, containing a paralel beta sheet
The handedness of b-a-b motif • Theoretically, two distinct “hands” can exist in b-a-b motif, with a helice above or below the plane of beta sheet • In almost all cases the right handed motif exists R L
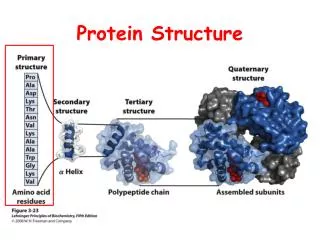
Domains • Domain ia a polypeptide chain or a part of a polypeptide chain that can fold indepedently in a stable tertiary structure with its own hydrophobic core • Domains can be formed from several simple motifs and additional secondary structure elements • Proteins can have anything from one to several tens of domains • In proteins with sevaral domains, most often each domain is associated with a distinct biological function
2xb hairpin + b strand 16xb-a-b 2x Greek key
C N C N C N • Domains are most often, but not always continuous pieces of primary structure
Example of proteins with several domains - lac repressor Helix-turn-helix domain (binds to DNA) hinge helix C-terminal helix (tetramerization) Core domain, containing two subdomains, which in turn contain several b-a-b motifs (binds ligand)
Intact IgG contains 12 immunoglobulin-like domains Each domain is made of two beta sheets with a topology similar to two Greek key motifs
The quaternary structure • Some proteins are biologically active as monomers. For those proteins quaternary structure does not exist • Other proteins, however, are active as homo- or hetero- polymers • The simplest case and by far the most common form of quaternary structure is a homodimer • The monomers in homopolymers are often arranged in a symmetric fashion with one or several symmetry axes going through the molecule or some sort of helical arrangement • Some biologically active units have a very complicated quaternary structure –like ribosomes or viral capsids
9-fold symmetry in light-harvesting complex II from Rhodopseudomonas acidophila.