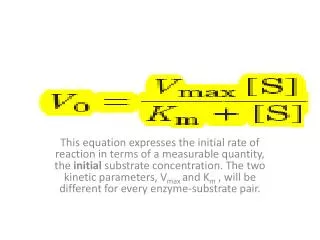Download

1 / 11
140 likes | 791 Views
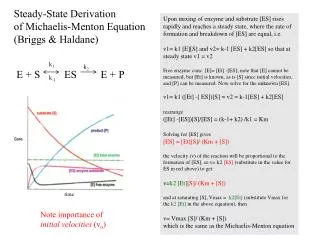
Prerequisits for Michaelis-Menten equation. 1.) refers to E + S ↔ ES → E + P equation where there is one substrate, and product conversion to ES is negligable 2.) steady state condition: [ ES ] is constant, so ES formation and degradation

E N D
Prerequisits for Michaelis-Menten equation 1.) refers to E + S ↔ ES → E + P equation where there is one substrate, and product conversion to ES is negligable 2.) steady state condition: [ES] is constant, so ES formation and degradation velocity are equal, appearance of P is linear in time, E is saturated with S 3.) slowest step is the formation of P, its rate constant is k2 4.) refers to initial velocity: [P]< 5% of [S], product does not compete with substrate 5.) [S]>>[E] (10-12 < [E ]< 10-7, 10-6 < [S] < 10-2 M)
Any reaction to proceed spontaneously needs net energy liberation: ΔG < 0. The speed of a reaction depends on the amount of the activation energy (Ea), the energetic barrier to be overcome. Arrhenius equation: rate constant = k = Ae-Ea/RT Enzymes catalyze the chemical reactions by decreasing the activation energy. They speed up the rate by creating a new route, the so called ES complex. Enzymes do not change the Gibbs free energy, but in case of themselves endothermic reactions, the energetically unfavourable reaction is coupled to an exothermic rection in spce and time, consequently the net reaction is negative. Enzymes bind the substrate to the substrate binding site by induced fit, so proper arrangement of active site groups fit only for the transition state of the ES complex. Multiple week interactions result strain in ES complex to accelerate the conversion to product.
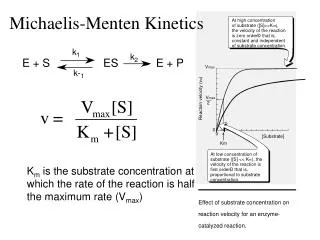
Characteristics of M-M kinetics 1.) Initial velocity / substrate cc. graph is right rectangular hyperbolic curve, assimptotes are vmax and –KM, Maximal velocity is reached at infinite S cc. 2.) If total enzyme cc.↑ then maximal velocity ↑ 3.) Michaelis-Menten constant is a constant in case of an E-S pair and pH, T, ionic strengh, other enzyme ligands are constant 4.) vmax depends on: pH, T, ionic strengh and type, presence of proper metal ions, coenzymes, activatory and inhibitory allosteric ligands, covalent modification of the enzyme, and enzyme amount 5.) double reciprocal 1/v versus 1/S curve is linear, intersepts: 1/vmax and 1/-KM 6.) Km = S cc. if v = vmax/2 depends on pH, T, ionic milieau
7.) Km = sum of ES degradation reaction velocity rate constants divided by ES formation reaction velocity rate constant k1 + k2 KM = ——— k-1 Michaelis-Menten equation: vi [S]—— = ———— hyperbolic curve: shaft of initial velocity curve approachesvmax KM + [S] but only reaches the maximal velocity at infinite S conc.
Why determine KM? • Physiological substrate concentration is around KM. • Measuring KM os different origin (species, tissues, cell organell) enzymes, we can determinetheir identity or difference (another protein). • Too big difference of assumed physiological value and in vitro value of KM means that anzymeactivator or inhibitor effects in the cell. • vmax can be measured, where [S] >> KM. • One enzyme can accept several substrates. The best S is that shich has the highest vmax/KMratio (meaning probably the lowest KM.) KM is increased bya.) a competitive inhibitor that competes with the S to bind to the active site orb.) a K-type allosteric inhibitor that when it binds to the allosteric binding site (not to the active site) decreases the enzyme affinity to substrate (less S will be bound). KM is decreased byan allosteric K-type activator that increases the affinity of the enzyme for substrate.
Effects of different parameters 1.) Reaction / incubation time Initial velocity should be: P(t) graph must be linear, time has to be short enough. If incubation time ↑, time ↑ for denaturation because of high temperature or pH 2.) pH: optimum depends on the amino acid composition of the enzyme proper ionization is necessary for S – catalytic site interaction; altered (rather extreme) pH leads to dehydration and denaturation of enzyme protein, the tertiary structure is disrupted. Different enzymes have different pH optimum peak and range. Blood: 7.4; stomach: 1-2; lysosomes, vesicles: 5; small intestine: 7-8 pH occurs.
3.) Temperature: if T increases, then number of effective collision of molecules in proper orientation will elevate, more molecules will have the activation energy, the sufficient energy for reaction. Velocity is increased at low T when T is increasing. If T increase, then kinetic energy of atoms in molecules will increase, bonds are disrupted leading to denaturation and lowering the volocity. The two opposite effect yealds the optimum curve, the top is around body temp. Together: temperature optimum depends on the interaction between amino acids
In steady state: E1 E2 E3 10.01 100 10.1 S → A → B → P ← ← ← 0.01 90 0.1 Forward velocities Reverse velocities 10.01 100 10.1 - 0.01 - 90 - 0.1 10 10 10 is the flux in the whole pathway Non-equilibrium near equilibrium non-equilibrium Irreversible reversible irreversible Rate-limiting = flux-generating
Near-equilibrium reaction Non-equilibrium reaction vf >> vr Vf≈ vr Reason: this enzyme activity and/or qantity is the smallest of all This enzyme is highly active compared to non-equilibrium enzymes Therefore: [A]≈[B] Therefore: [S]>>[A] If [S]↑, then vf not↑ because E1 is saturated with S and v = vmax If [A]↑, then A → B and vf↑ E2 is not saturated with A v <<vmax If [A]↑, then vr↑, but [S]↑ is neglectable If [B]↑,then A ← B and vr↑, [A] ↑ [B[eq [A]eq [B]real [A]real [A]eq [S]eq [A]real [S]real ≈ >> Keq >>Q Keq ≈ Q
Keq Q ΔG = ΔG°´ + RT lnQ = -RT lnKeq + RT lnQ = - RT ln Non-equilibrium reaction near-equilibrium reaction ΔG << 0 ΔG ≈ 0 If E2 conc. or activity is increased, then both vf and vr will elevate, therefore equilibrium is not changed If E1 concentration or activity is increased, then vmax will elevate, and product „A” grows up E1 activity can be regulated by allostericaly and/ or covalent modification E2 is regulated by substrate and/or product concentration change E1 concentration can be changed at transcription or translation level E2 is not regulated by ligands or other way
Flux-generating reaction Is that non-equilibrium reaction in the beginning of a pathway that is saturated by the pathway substrate and determines the speed of the whole pathway, ie. each reactions in the pathway has the same velocity. A metabolic pathway is a series of enzyme-catalized reactions, initiated by a flux-generating step and ending with either the loss of products to the environment, to a stored product (a metabolic ‘sink’) or in a reaction that precedes another flux-generating step (that is the beginning of the next pathway.