Download
1 / 1
10 likes | 168 Views
#___. Genetic Correlates of Phenotypic Hypersusceptibility to Efavirenz (EFV) Among 444 Baseline Isolates from Five ACTG Studies N Shulman*, R Bosch, J Mellors, M Albrecht, and D Katzenstein DACS 217 Study Team, NIAID Sponsored AIDS Clinical Trials Group, Bethesda, MD. *Nancy Shulman, MD

E N D
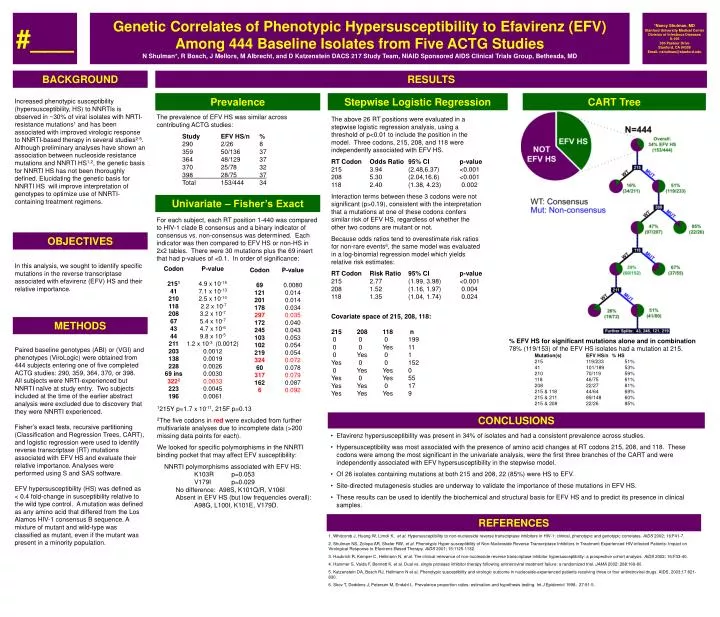
#___ Genetic Correlates of Phenotypic Hypersusceptibility to Efavirenz (EFV) Among 444 Baseline Isolates from Five ACTG Studies N Shulman*, R Bosch, J Mellors, M Albrecht, and D Katzenstein DACS 217 Study Team, NIAID Sponsored AIDS Clinical Trials Group, Bethesda, MD *Nancy Shulman, MD Stanford University Medical Center Division of Infectious Diseases S-156 300 Pasteur Drive Stanford, CA 94305 Email: nshulman@stanford.edu BACKGROUND RESULTS Prevalence Stepwise Logistic Regression CART Tree Increased phenotypic susceptibility (hypersusceptibility, HS) to NNRTIs is observed in ~30% of viral isolates with NRTI-resistance mutations1 and has been associated with improved virologic response to NNRTI-based therapy in several studies2-5. Although preliminary analyses have shown an association between nucleoside resistance mutations and NNRTI HS1,2, the genetic basis for NNRTI HS has not been thoroughly defined. Elucidating the genetic basis for NNRTI HS will improve interpretation of genotypes to optimize use of NNRTI-containing treatment regimens. The prevalence of EFV HS was similar across contributing ACTG studies: Study EFV HS/n % 290 2/26 8 359 50/136 37 364 48/129 37 370 25/78 32 398 28/75 37 Total 153/444 34 The above 26 RT positions were evaluated in a stepwise logistic regression analysis, using a threshold of p<0.01 to include the position in the model. Three codons, 215, 208, and 118 were independently associated with EFV HS. RT Codon Odds Ratio 95% CI p-value 215 3.94 (2.48,6.37) <0.001 208 5.30 (2.04,16.6) <0.001 118 2.40 (1.38, 4.23) 0.002 Interaction terms between these 3 codons were not significant (p>0.19), consistent with the interpretation that a mutations at one of these codons confers similar risk of EFV HS, regardless of whether the other two codons are mutant or not. Because odds ratios tend to overestimate risk ratios for non-rare events6, the same model was evaluated in a log-binomial regression model which yields relative risk estimates: RT Codon Risk Ratio 95% CI p-value 215 2.77 (1.99, 3.98) <0.001 208 1.52 (1.16, 1.97) 0.004 118 1.35 (1.04, 1.74) 0.024 Univariate – Fisher’s Exact For each subject, each RT position 1-440 was compared to HIV-1 clade B consensus and a binary indicator of consensus vs. non-consensus was determined. Each indicator was then compared to EFV HS or non-HS in 2x2 tables. There were 30 mutations plus the 69 insert that had p-values of <0.1. In order of significance: OBJECTIVES In this analysis, we sought to identify specific mutations in the reverse transcriptase associated with efavirenz (EFV) HS and their relative importance. Codon 2151 41 210 118 208 67 43 44 211 203 138 228 69 ins 3222 223 196 P-value 4.9 x 10-15 7.1 x 10-13 2.5 x 10-10 2.2 x 10-7 3.2 x 10-7 5.4 x 10-7 4.7 x 10-6 9.8 x 10-5 1.2 x 10-3 (0.0012) 0.0012 0.0019 0.0026 0.0030 0.0033 0.0045 0.0061 Codon 69 121 201 178 297 172 245 103 102 219 324 60 317 162 6 P-value 0.0080 0.014 0.014 0.034 0.035 0.040 0.043 0.053 0.054 0.054 0.072 0.078 0.079 0.087 0.092 Covariate space of 215, 208, 118: 215 208 118 n 0 0 0 199 0 0 Yes 11 0 Yes 0 1 Yes 0 0 152 0 Yes Yes 0 Yes 0 Yes 55 Yes Yes 0 17 Yes Yes Yes 9 METHODS • % EFV HS for significant mutations alone and in combination • 78% (119/153) of the EFV HS isolates had a mutation at 215. • Mutation(s) EFV HS/n % HS • 215 119/233 51% • 41 101/189 53% • 210 70/119 59% • 118 46/75 61% • 208 22/27 81% • 215 & 118 44/64 69% • 215 & 211 89/148 60% • 215 & 208 22/26 85% Paired baseline genotypes (ABI) or (VGI) and phenotypes (ViroLogic) were obtained from 444 subjects entering one of five completed ACTG studies: 290, 359, 364, 370, or 398. All subjects were NRTI-experienced but NNRTI naïve at study entry. Two subjects included at the time of the earlier abstract analysis were excluded due to discovery that they were NNRTI experienced. Fisher’s exact tests, recursive partitioning (Classification and Regression Trees, CART), and logistic regression were used to identify reverse transcriptase (RT) mutations associated with EFV HS and evaluate their relative importance. Analyses were performed using S and SAS software. EFV hypersusceptibility (HS) was defined as < 0.4 fold-change in susceptibility relative to the wild type control. A mutation was defined as any amino acid that differed from the Los Alamos HIV-1 consensus B sequence. A mixture of mutant and wild-type was classified as mutant, even if the mutant was present in a minority population. 1215Y p=1.7 x 10-11, 215F p=0.13 2The five codons in red were excluded from further multivariate analyses due to incomplete data (>200 missing data points for each). We looked for specific polymorphisms in the NNRTI binding pocket that may affect EFV susceptibility: NNRTI polymorphisms associated with EFV HS: K103R p=0.053 V179I p=0.029 No difference: A98S, K101Q/R, V106I Absent in EFV HS (but low frequencies overall): A98G, L100I, K101E, V179D. CONCLUSIONS • Efavirenz hypersusceptibility was present in 34% of isolates and had a consistent prevalence across studies. • Hypersusceptibility was most associated with the presence of amino acid changes at RT codons 215, 208, and 118. These codons were among the most significant in the univariate analysis, were the first three branches of the CART and were independently associated with EFV hypersusceptibility in the stepwise model. • Of 26 isolates containing mutations at both 215 and 208, 22 (85%) were HS to EFV. • Site-directed mutagenesis studies are underway to validate the importance of these mutations in EFV HS. • These results can be used to identify the biochemical and structural basis for EFV HS and to predict its presence in clinical samples. REFERENCES 1. Whitcomb J, Huang W, Limoli K, et al. Hypersusceptibility to non-nucleoside reverse transcriptase inhibitors in HIV-1: clinical, phenotypic and genotypic correlates. AIDS 2002; 16:F41-7. 2. Shulman NS, Zolopa AR, Shafer RW, et al. Phenotypic Hyper-susceptibility of Non-Nucleoside Reverse Transcriptase Inhibitors in Treatment Experienced HIV-infected Patients: Impact on Virological Response to Efavirenz-Based Therapy. AIDS 2001; 15:1125-1132. 3. Haubrich R, Kemper C, Hellmann N, et al. The clinical relevance of non-nucleoside reverse transcriptase inhibitor hypersusceptibility: a prospective cohort analysis. AIDS 2002; 16:F33-40. 4. Hammer S, Vaida F, Bennett K, et al. Dual vs. single protease inhibitor therapy following antiretroviral treatment failure: a randomized trial. JAMA 2002; 288:169-80. 5. Katzenstein DA, Bosch RJ, Hellmann N et al. Phenotypic susceptibility and virologic outcome in nucleoside-experienced patients receiving three or four antiretroviral drugs. AIDS, 2003;17:821-830. 6. Skov T, Deddens J, Petersen M, Endahl L. Prevalence proportion ratios: estimation and hypothesis testing. Int J Epidemiol 1998; 27:91-5.




















