Download
1 / 39
400 likes | 664 Views
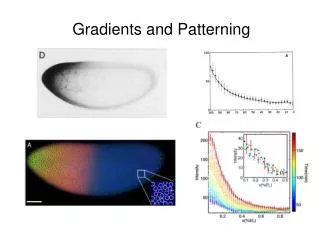
Gradients and Patterning. Gradients and Patterning. Positional Information: the means by which a cell can “sense” its position within an embryo and act accordingly Morphogen: a factor that controls cell identity by acting at a distance and in a concentration-dependent manner

E N D
Gradients and Patterning Positional Information: the means by which a cell can “sense” its position within an embryo and act accordingly Morphogen: a factor that controls cell identity by acting at a distance and in a concentration-dependent manner (different concentrations= different identities) Organizer: a group of cells or tissue that patterns surrounding cell (usually by secreting a morphogen) (Lewis Wolpert, 1969)
Test of a Morphogen Does a signal act in a concentration-dependent manner to specify DIFFERENT cell identities? Some Alternatives: 1) The “relay” signal Signal specifies “blue” cells, which then use different signal to make white cells 2) The “permissive” signal The field of cells has already been patterned (prepattern), and the signal just tells the cells to act (e.g. regulates timing) 3) Cells all originated close to signal, and then moved away due to proliferation or cell migration. Different identities could be specified by timing.
Some “Performance Objectives” of Morphogen Gradients: Lander PMID: 17254964 Multiple responses (cell identities) to one signal Sharp boundaries of responses -pure diffusion gradients are “shallow” -need additional control of gradient or response Timescale -diffusion gradients can take hours to reach steady-state -many patterning events occur much faster Robustness -environmental or genetic conditions can change -patterning events need to resist change Scaling -field size changes over time or between related species -morphogen gradients need to scale reponses accordingly
The Maternal-Effect Mutants (also found the D/V Group) Nusslein-Volhard, 2004
bicoid mRNA oskar mRNA protein + nanos RNA microtubule-based - + A-P polarity set up in egg chamber
bicoid is the Anterior Determinant After Frigerio, 1986 Berleth, 1988 Driever, Seigel and Nusslein-Volhard, 1990
bcd dose 1X 2X 3X 4X
Live Imaging and Quantitation of the BCD Gradient Gregor et al., Cell 2007a and b
The Synthesis, Diffusion and Degradation Model (SDD) -BCD synthesized from anteriorly localized RNA -Diffuses through embryo to form gradient -Degradation of protein determines lifetime Variations of SDD -Steady-State Model: BCD lifetime is short, so gradient reaches a stabel steady-state when it is “read out” by cells -Pre-Steady-State Model: BCD lifetime is longer, so gradient is still changing when it is “read out”. In this case, WHEN it is read is critical for the final pattern -Nuclear Trapping Model: Since the number of nuclei goes from 1 to 6000 during patterning, BCD nuclear transport and DNA binding can dramatically affect gradient Problem: the measured Diffusion Coefficient for BCD is 10 TIMES to small to account for gradient formation by SDD in relevant timescale!
The bcd RNA also moves posteriorly and may be actively transported
“Old” diffusion coefficient (FRAP) = 0.3 µm2/sec FCS calculated diffusion coefficient = 7 µm2/sec
Scaling of the BCD Gradient in Different Size Embryos L. sericata D. melanogaster BCD Gradient scales to size of embryo so gradient is similar relative to A/P position L. sericata D. melanogaster PMID:16352710 eGFP-BCD from different size species gives similar gradient in D. melanogaster Therefore difference in length constant NOT function of protein D. melanogaster D. busckii
To Understand and Model BCD Gradient We (STILL) Need to Know: Where is BCD produced? Is RNA or translation localized to anterior at critical window or is it an RNA gradient? Is production rate constant? Models about gradient length and stability ASSUME constant translation, but this has not been measured. How does BCD move? Is there a “fast” species at 3.0 µm2/sec? Is BCD transported or does it just diffuse? Is the RNA transported? How is BCD distributed between different compartments? Cytoplasmic, nuclear, DNA bound, cytoskeletal bound? What is BCD’s lifetime? Short lifetime favors reaching steady state and a stable gradient. Long lifetime means more likely that gradient is constantly changing, meaning it needs to be “read out” at correct time. See PMID: 20570935
Shallow gradients can make sharp boundaries PMID: 18818726
1996 Footprinting of A1 site in context of: Intact hb enhancer Hill coefficient = 5 A1 + X1 sites only A1 site alone
A Single Gradient Can Generate Multiple Domains of Gene Expression -Even a shallow gradient can result in sharp boundaries of expression -Different promoters can respond to different [Bcd] Crauk and Dostatni, 2005
Complex Patterns From Simple Ones: Gap Genes BCD Giant Kruppel
BCD is Also a Translation Factor Reviewed by Ephrussi and St. Johnston, 2004
Extracellular Signaling Pathways: More Common Morphogens A finite number of ligands and signal transduction pathways must create much of the cellular diversity seen in a developing embryo. The same pathways can generate different outputs by: -Creating different responses at different concentrations (Morphogens) -Causing different responses depending on cellular history (i.e. a cell’s prior identity when it sees the signal affects how it responds) -Pathways being used in combination Some relevant pathways for signaling cell identity: hedgehog wingless (wnt) TGF-ß/BMP/Activin Notch Toll Tyrosine kinase receptors (EGF-R, FGF-R, etc.) Jak/Stat Toll/IL Tor G Protein Coupled Receptors (peptide and non-peptide ligands Nuclear Hormone Receptors
Ligand Production -synthesis -modification -secretion Ligand Distribution Ligand “Activity” Receptor -distribution -co-receptors Signaling -transduction -downregulation Output -transcriptional regulation -other outputs
Shaping Morphogen Gradients Regulating Extracellular Ligands: Ligand production: “Source” -Expression (transcription and translation) -Protein Processing (proligand cleavage) -Protein Modification (e.g. addition of lipids)
Autoproteolysis and Lipid Modification of Hh skinny hh Beachy Lab
Shaping Morphogen Gradients Regulating Extracellular Ligands: Ligand production: “Source” -Expression (transcription and translation) -Protein Processing (proligand cleavage) -Protein Modification (e.g. addition of lipids) Extracellular Regulators -Inhibitors -Carriers -Ligand Heterodimers
Dpp is a Morphogen for the Dorsal Ectoderm DPP D V SOG
eve st2-GFP-DPP RNA wt Extracellular Protein (PVI) sog pMAD Extracellular Protein (PVI) Extracellular DPP (PVI) D A Gradient of DPP Protein Forms From Broadly Expressed Dpp RNA Side view V Dpp RNA Dorsal view O’Connor and Ferguson Labs
Extracellular Regulators of Dpp -Short gastrulation (Sog) and Twisted Gastrulation (Tsg) bind and inhibit Dpp -They also are extracellular “carriers” for Dpp -Sog is only expressed in ventral regions -Tolloid (Tld) is a protease that degrades Sog when Sog is bound to Dpp -Screw is a second BMP-type ligand (like Dpp) that can form heterodimers with Dpp -The Dpp/Screw heterodimer is a more potent signaling complex -Sog preferrentially binds the Dpp/Screw heterodimer
Extracellular Regulators of Dpp Shimmi et al., Cell 2005
Shaping Morphogen Gradients Regulating Extracellular Ligands: Ligand production: “Source” -Expression (transcription and translation) -Protein Processing (proligand cleavage) -Protein Modification (e.g. addition of lipids) Extracellular Regulators -Inhibitors -Carriers -Ligand Heterodimers 3) Ligand movement: Do morphogens ever diffuse? -Modifications affect ligand distribution -Interactions with extracellular proteins (eg. Heparan sulfate proteoglycans) -Movement within intracellular vessicles (transcytosis) or extracellular vessicles (argosomes/exosomes) -Cell movements and proliferation: displacement of cells away from a signaling source can create a gradient of signaling (in time or space)
Models of Gradient Formation Diffusion S. Cohen Transcytosis Gonzales-Gaitan Cell 2000 GFP-DPP DPP Target shi clone
Do ligands remain embedded in membranes (“argosomes”)? Or, could the cell go to the signal, rather than the signal to the cell? Eaton Lab Cytonemes Kornberg Lab
fgf8 is only transcribed in the tailbud A model Morphogen diffusion: RNA Inheritence: Exon Probe Intron Probe Proliferation + Decay = Gradient Fgf8 Gradient Formation by RNA Inheritance New transcription is not required for gradient formation
Shaping Morphogen Gradients Regulating Extracellular Ligands: Ligand production: “Source” -Expression (transcription and translation) -Protein Processing (proligand cleavage) -Protein Modification (e.g. addition of lipids) Extracellular Regulators -Inhibitors -Carriers -Ligand Heterodimers 3) Ligand movement -Models for ligand movement: Do morphogens ever diffuse? -Modifications affect ligand distribution -Interactions with extracellular proteins (eg. Heparan sulfate proteoglycans) -Cell movements and proliferation: displacement of cells away from a signaling source can create a gradient of signaling (in time or space) 4) Ligand degradation: “Sink” -Extracellular degradation -Receptor binding can titrate ligand -Endocytosis (involved in ligand signaling and degradation as well as possibly movement)
wg-expressing Increased wg degradation wg-HRP fusion protein Regulated Endocytosis and Degradation Shapes the Wg Gradient Dubois et al. Cell 2001
Endocytosis regulates signaling in multiple ways 1) Endocytosis facilitates movement of the Dpp ligand Endocytosis blocked in posterior Range of Dpp ligand reduced 2) Signal Transduction: Activation of Smad facilitated by SARA in clathrin-dependent vesicles 3) Signal Downregulation: Degradation of receptors in caveolin-containing vesicles Gonzales-Gaitan, 2003