Download

1 / 52
560 likes | 1.16k Views
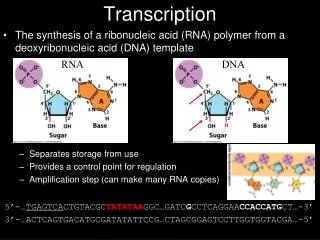
Mechanisms of Transcription. RNA polymerase does not need a primer to initiate transcription. The RNA product does not remain base-paired to the template DNA strand. Transcription is less accurate than replication. RNA polymerases and the transcription cycle.

E N D
RNA polymerase does not need a primer to initiate transcription. • The RNA product does not remain base-paired to the template DNA strand. • Transcription is less accurate than replication.
RNA polymerases and the transcription cycle • RNA polymerase performs essentially the same reaction in all cells. • From bacteria to mammals, the cellular RNA polymerases are made up of multiple subunits.
Transcription is catalyzed by RNA polymerase, which catalyzes the synthesis of RNA using DNA as a template. RNA polymerase from E.coli contains two subunits,’, , ω and . The E. coli holoenzyme is composed of a core, which is competent to carry out RNA synthesis, and a σ factor, which directs the core to transcribe specific genes.
Comparison of the crystal structures of prokaryotic and eukaryotic RNA polymerases
Transcription in Prokaryotic cells Transcription involves three stages
Initiation: A promoter is the DNA sequence that initially binds the RNA polymerase. Only one of the DNA strands acts as a template. The choice of promoter determines which stretch of DNA is transcribed and is the main step at which regulation is imposed. • Elongation: Once the RNA polymerase has synthesized a short stretch of RNA approximately ten bases), it shifts into the elongation phase. • Termination: Once the polymerase has transcribed the length of the gene, it must stop and release the RNA product.
The transcription cycle in bacteria • Bacterial promoters vary in strength and sequence, but have certain defining features • 3 regions of conservation: -35, -10 and the length of spacer 17 - 19 bp, called consensus sequence. • Promoters with sequences closer to the consensus sequence are generally stronger than those that match less well.
An additional DNA element, UP-element, that binds RNA polymerase is found in some strong promoters and increase polymerase binding by providing an additional specific interaction between the enzyme and the DNA. • Another class of promoters lacks a -35 region and instead has a so-called “ extended -10” element.
The sigma factors mediates binding of polymerase to the promoter
The best evidence for the functions of these regions shows that sub-regions 2.4 and 4.2 are involved in promoter –10 box and –35 box recognition. The extended –10 element is recognized by an αhelix in σregion 3. σ70 family: There are four conserved regions in sigma 70 family proteins.
The RNA polymerase α subunit has an independently folded C-terminal domain that can recognize and bind to a promoter’s UP element. This allows very tight binding between polymerase and promoter. The α -CTD is connected to the αNTD by a flexible linker. α subunit response to activator, repressor, elongation factor and transcription factors
Transcription Initiation involves three defined steps • The first step in RNA synthesis is the binding of RNA polymerase to a DNA promoter- to form what is called a closed complex. • In the second step, the closed complex undergoes a transition to the open complex in which the DNA strands separate over a distance of some 14 bp around the start site.
Once an RNA polymerase molecule has bound to a promoter site and locally unwound the DNA double helix, initiation of RNA synthesis can take place. • Once the enzyme gets further than 10 bp, it is said to have escaped the promoter. A stable ternary complex contains enzyme, DNA and RNA. This is a transition to elongation phase.
Transition to the open complex involves structural changes in RNA polymerase and in the promoter DNA
The transition from closed to open complex involves structural changes in the enzyme and the opening of the DNA double helix to reveal the template and nontemplate strands. In bacterial enzyme with σ70, this transition called isomerization, does not require energy from ATP hydrolysis. The active site of the enzyme, which is made up of regions from both the β and β’ subunits, is at the base of the pincers within the active center cleft.
Transcription is initiated by RNA polymerase without the need for a primer • This requires that the initiating ribonucleotide be brought into the active site and held stably on the template while the next NTP is presented with correct geometry for the polymerization to occur. • The enzyme has to make specific interactions with the initiating NTP. • The interactions are specific for the nucleotide on A, and only chains initiated with A are held in a manner suitable for efficient initiation.
RNA polymerase synthesizes several short RNAs before entering the elongation phase • Abortive initiation: the enzyme synthesizes short RNA molecules of less than ten nucleotides and then released from the polymerase. And the enzyme begins RNA synthesis again. • Once a polymerase manages to make an RNA longer than 10 bp, a stable ternary complex is formed. This is the start of the elongation phase. • Region 3.2 of σ factor may be involved, and it mimics RNA.
The elongation polymerase is a processive machine that synthesizes and proofreading RNA • Double-stranded DNA enters the front of the enzyme between the pincers. • At the opening of the catalytic cleft, the strands separate to follow different paths through the enzyme before exiting via their respective channels and reforming a double helix behind the elongation polymerase.
RNA polymerase carries out two proofreading functions • Pyrophosphorolytic editing: the enzyme uses its active site to catalyze the removal of an incorrectly inserted NTP. • Hydrolytic editing: the polymerase backtracks by one or more nucleotides and cleaves the RNA product, removing the error-containing sequence. • Hydrolytic editing is stimulated by Gre factors, which also serves as elongation stimulating factors.
Transcription is terminated by signals within the RNA sequences • In bacteria, terminators come in two types: rho-independent and rho-dependent.
Rho-independent Terminators, also called intrinsic terminators, a short inverted repeats (about 20 nucleotides) followed by a string of about eight A:T bp.
Rho-dependent terminators • Have less well-characterized RNA elements and requires the action of the rho factor. • Rho is an RNA helicase, composed of 6 identical subunits, each subunit has an RNA binding domain and ATPase domain, requires ATP to function. • Rho releases the RNA product from the DNA template.
How is Rho directed to a particular RNA molecule? • There is some specificity in the sites it binds (the rut sites, Rho utilization). Optimally these sites consist of stretches of about 40 nucleotides that do not fold into a secondary structure; they are also rich in C. • Rho fails to bind any transcript that is being translated.
Transcription in Eukaryotes • Eukaryotic cells have three different polymerase (Pol I, II and III) • Whereas bacteria require only one initiation factor, several initiation factors are required for efficient and promoter-specific initiation in eukaryotes. These are called the general transcription factors (GTFs).
Three different RNA polymerases • RNA polymerase I resides in the nucleolus and is responsible for synthesizing three of the four types of rRNA found in eukaryotic ribosomes (28S, 18S,and 5.8 S rRNA). • RNA polymerase II is found in the nucleoplasm and synthesizes precursors to mRNA, the class of RNA molecules that code for proteins. • RNA polymerase III is also a nucleoplasmic enzyme, but it synthesizes a variety of small RNAs, including tRNA precursors and the smallest type of ribosomal RNA, 5S rRNA.
Core promoter: defined as the minimal set of DNA sequences sufficient to direct the accurate initiation of transcription by RNA polymerase Four types of DNA sequences are involved in core promoter function in RNA polymerase II (1) a short initiator (Inr) (2) the TATA box (3) the TFIIB recognition element (BRE) (4) the down stream promoter element (DPE) RNA polymerase II core promoters are made up of combinations of four different sequence elements
Regulatory sequences • Beyond the core promoter, there are other sequence elements required for efficient transcription in vivo. • Together these elements constitute the regulatory sequences: promoter proximal elements, upstream activator sequences, enhancers, silencers, boundary elements and insulators. All these elements bind regulatory proteins.
RNA polymerase II forms a pre-initiation complex with general transcription factors at the promoter • The complete set of general transcription factors and polymerase, bound together at the promoter and poised for initiation, is called the pre-initiation complex.
In vitro, TFIIA, TFIIB, TFIIF together with polymerase, and then TFIIE and TFIIH form the pre-initiation complex. Promoter melting in eukaryotes requires hydrolysis of ATP and is mediated by TFIIH. In eukaryotes, promoter escape involves phosphorylation of the polymerase.
The largest subunit of Pol II has a C-terminal domain (CTD), which extends as a tail. The CTD contains a series of repeats of the heptapeptide sequence: Tyr-Ser-Pro-Thr-Ser-Pro-Ser. They are 27 of these repeats in the yeast and 52 in the human case. • Each repeat contains sites for phosphorylation by specific kinases including that is a subunit of TFIIH.
TBP binds to and distorts DNA using a beta sheet inserted into the minor groove TBP uses an extensive region of sheet to recognize the minor groove of the TATA element. It also bends the DNA by an angle of approximately 80o The interaction between TBP and DNA involves only a limited number of hydrogen bonds between the protein and the edges of the base pairs in the minor groove. TBP-DNA complex
The other General Transcription factors also have specific roles in initiation • TAFs: TBP is associated with about ten TAFs. • TFIIB: a single polypeptide chain, enters the pre-initiation complex after TBP. This protein appears to bridge the TATA-bound TBP and polymerase. • TFIIF: This two-subunit factor associated with PolII and is recruited to the promoter together with the enzyme.
TFIIE and TFIIH:TFIIE recruits and regulates TFIIH. • TFIIH controls the ATP-dependent transition of the pre-initiation complex to the open complex. It has nine subunits. Within TFIIH are two subunits that function as ATPases, and another that is a protein kinase, with roles in promoter melting and escape
In vivo, transcription Initiation requires additional Proteins, including the mediator complex • The high, regulated levels of transcription in vivo require the Mediator Complex, transcriptional regulatory proteins and in many cases, nucleosome-modifying enzymes. • Mediator is associated with the CTD “tail” of the large polymerase subunit through one surface, while presenting other surfaces for interaction with DNA-bound activators.
Different Mediator subunits to bring polymerase to different genes. • Mediator aids initiation by regulating the CTD kinase in TFIIH. • The need for nucleosome modifiers and remodellers also differs at different promoters.
Mediator consists of many subunits, some conserved from yeast to human • There are various forms of Mediator, each containing subsets of Mediator subunits. • A complex consisting of Pol II, Mediator, and a some of the general transcription factors can be isolated from cells as a single complex in the absence of DNA---RNA Pol II holoenzyme.
A new set of factors stimulate Pol II elongation and RNA proofreading • Elongation factors (such as TFIIS and hSPT5) are factors stimulate elongation. • Phosphorylation of the CTD leads to an exchange of initiation factors for those factors required for elongation and RNA processing. • Various proteins are thought to stimulate elongation by Pol II. One of them is the kinase P-TEFb.
TFIIS • TFIIS stimulates the overall rate of elongation by limiting the length of time polymerase pauses. • TFIIS stimulates an inherent RNAse activity in polymerase and contributes to proofreading by polymerase.
Kinase P-TEFb • Kinase P-TEFb, is recruited to polymerase by transcriptional activators. • Once bound to Pol II, it phosphorylates the serine residue at position 2 of the CTD. • P-TEFb phosphorylates and activates hSPT5, another elongation factor. hSpT5 stimulates 5’ capping enzyme. • P-TEFb also recruits TAT-SF1, an elongation factor, to stimulate elongation. TAT-SF1 recruits components of the splicing machinery.
Elongating Polymerase is associated with a new set of protein factors required for various types of RNA Processing • There is an overlap in proteins involved in elongation, and those required for RNA processing. • hSPT5 recruits and stimulates the 5’ capping enzyme. Elongation factor TAT-SF1 recruits components of the splicing machinery. Phosphorylation on ser 5: capping factors Phosphorylation on ser 2: splicing factors
The structure and formation of the 5’ RNA cap • The first RNA processing event is capping. • It is a methylated guanine, and it is joined to the RNA transcript by an unusual 5’ -5’ linkage. • The RNA is capped when it is still only 20-40 nucleotides long- when the transcription cycles has progressed only to the transition between the initiation and elongation phases.
Polyadenylation and termination • The final RNA processing event, polyadenylation of the 3’ end of the mRNA, is linked with the termination of trancription. • The polymerase CTD tail is involved in recruiting the enzymes necessary for polyadenylation. • Once polymerase has reached the end of a gene, it encounters specific sequences called poly-A signals.
CPSF (cleavage and polyadenylation specificity factor) and CstF (cleavage stimulation factor) are carried by the CTD of polymerase as it approaches the end of the gene. • Once the CPSF and CstF are bound to the RNA, other proteins are recruited as well, leading initially to RNA cleavage and then polyadenylation. • Polyadenylation signal is required for termination.
RNA polymerase I and III recognize distinct promoters, using distinct sets of transcription factors, but still require TBP • Each of these enzymes also works with its own unique set of general transcription factors. However, TBP is universal for most of the cases.