Download

1 / 63
650 likes | 1.07k Views
P entoso fosfátová dráha. Metabolismus fruktosy a galaktosy. Vznik glukuronové kyseliny a aminocukrů. Biochemick ý ústav LF MU 2012 (E.T.). Pentosofosfátová dráha. Tk áňová lokalizace:

E N D
Pentosofosfátová dráha. Metabolismus fruktosy a galaktosy. Vznik glukuronové kyseliny a aminocukrů. Biochemický ústav LF MU 2012 (E.T.)
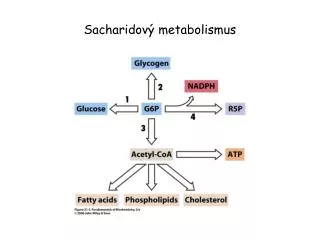
Pentosofosfátová dráha Tkáňová lokalizace: ve velkém rozsahu játra, tuková tkáň (až 50% metab.glukosy), erytrocyty, štítná žláza, laktující mléčná žláza ad. (obecně tkáně, kde probíhají redukční syntézy) Ostatní tkáně využívají jen některé reakce Buněčná lokalizace: cytoplasma
Význam pentosofosfátové dráhy • zdroj NADPH (redukční syntézy, oxygenasy se smíšenou funkcí, redukce glutathionu) • zdroj ribosa-5-P (nukleové kyseliny, nukleotidy) • zapojení pentos přijatých potravou do metabolismu Neslouží k zisku energie, ani energii přímo nespotřebovává
Dvě části pentosofosfátové dráhy oxidační část nevratné reakce neoxidační část (regenerační) vratné reakce
Oxidační část pentosofosfátové dráhy NADP+ NADPH + H+ 6-fosfoglukonolakton Glukosa-6-P laktonasa glukosa-6-P-dehydrogenasa 6-fosfoglukonát NADP+ NADPH + H+ 6-fosfoglukonátdehydrogenasa Ribulosa-5-P + CO2 Regulace: inhibice NADPH (inhibice produktem)
O C O - H C O H H O C H C O H H H C O H H C O P H Oxidační část pentosofosfátové dráhy (podrobněji) – vznik 6-fosfoglukonátu NADPH + H+ NADP+ H2O C H O P C H O P 2 2 O O O H laktonasa O H glukosa-6-P-dehydrogenasa O H O H O H O H O H O H 6-fosfoglukono--lakton Glukosa-6-P 6-fosfoglukonát
Oxidační část pentosofosfátové dráhy(podrobněji) –přeměna 6-fosfoglukonátu O C O - H H C O H H C O H H O C H C O H C O H H C O H H C O H H C O H H C O P H C O P H H NADP+ NADPH + H+ 6-phosphoglukonátdehydrogenasa CO2 6-fosfoglukonát ribulosa-5-P Ziskem oxidační větve pentosového cyklu jsou 2 moly NADPH a pentosa fosfát
Regenerační fáze pentosofosfátové dráhy (pokud pentosy nejsou využity pro syntézu nukleotidů) souhrnná rovnice: 3 Ribulosa-5-P 2 Fruktosa-6-P + Glyceraldehyd-3-P Proč regenerace ? Některé buňky potřebují mnoho NADPH. Při jeho produkci vzniká velké množství pentos, které buňka nepotřebuje.
H H C O H C O C H O H H C O H H C O P H Enzymy v regenerační fázi pentosofosfátové dráhy Isomerasa Syntéza nukleotidů Ribosa-5-P Ribulosa-5-P Pentosový cyklus – regenerační fáze
H H C O H C O C H O H H C O H H C O P H Epimerasa Ribulosa-5-P Xylulosa-5-P
H H C O H C O H O C H H C O H H C O P H Transketolasa - přenáší dvouuhlíkatý zbytek H H C O H C O C H HO + H C O H + H C O H H C O H H C O P Glyceraldehyd-3-P H Sedoheptulosa-7-P Ribosa-5-P Xylulosa -5-P 5C 5C 3C 7C + + Kofaktor: thiamindifosfát
H H C O H C O O C H H C O H H C O H H C O H H H C O P H C O H H C O H O C H H C O H H C O H H C O P H Transaldolasa - přenáší tříuhlíkatý zbytek H + + Glyceraldehyd-3-P Erythrosa-4-P Sedoheptulosa-7-P Fruktosa-6-P + 4C + 6C 7C 3C
Transketolasa-přenáší dvouuhlíkatý zbytek H H C O H + C O + H O C H H C O H H C O H Erythrosa-4-P H C O P Xylulosa -5-P H Fruktosa-6-P Glyceraldehyd-3-P 3C 5C 6C + 4C +
Souhrnná bilance pentosafosfátové dráhy Ribulosa-5-P Ribosa -5-P 2 Ribulosa-5-P 2 Xylulosa -5-P Xylu-5-P + Rib-5-P Glyc-3-P + Sed-7-P Sed-7-P + Glyc-3-P Ery-4-P + Fru-6-P Xylu-5-P + Ery-4-P Glyc-3-P + Fru-6-P 3 Ribulosa-5-P Glyceraldehyd-3-P + 2 Fru-6-P 3C + 2 x 6C 3 x 5C
H H C O H C O O C H H C O H H C O H H C O H H C O P H H H C O H C O H O C H H C O H H C O H H C O P H H Erytrosa-4-P Xylulosa-5-P Ribosa-5-P TK TA TK H H C O H C O Ribulosa-5-P H O C H H C O H H C O P Xylulosa-5-P Glyceraldehyd-3-P H
Tvorba ribosafosfátu z meziproduktů glykolýzy Reakce regenerační fáze pentosového cyklu jsou vratné. To umožňuje produkovat pentosy v buňce i v situacích, kdy není potřebné NADPH, pouhým zvratem reakcí regenerační fáze, z meziproduktů glykolýzy.
Transketolasová reakce v opačném směru fruktosa-6-P + glyceraldehyd-3-P erytrosa-4-P + xylulosa-5-P (z glykolýzy) Transaldolasová reakce v opačném směru erytrosa-4-P + fruktosa-6-P sedoheptulosa-7-P + glyceraldehyd-3-P (z glykolýzy) Další transketolasová reakce v opačném směru sedoheptulosa-7-P + glyceraldehyd-3-P 2 pentosy
Regulace pentosofosfátové dráhy • rychlost závisí na úvodních nevratných reakcích oxidační fáze (viz též snímek 5) • dostupnost substrátu (NADP+), inhibice produktem • indukce enzymů insulinem
Reakce vyžadující NADPH • redukce oxidovaného glutathionu • monooxygenázové reakce s cytP450 • respirační vzplanutí v leukocytech • redukční syntézy: • syntéza mastných kyselin • elongace mastných kyselin • syntéza cholesterolu a steroidů • syntéza nukleotidů • syntéza NO z argininu
NADH x NADPH / srovnání * Transhydrogenasa v mitochondriální membráně může katalyzovat přenos H z NADH na NADP+
Význam pentosofosfátové dráhypro erytrocyty Pentosový cyklus je jediným zdrojem NADPH pro erc 5-10% glukosy v erc NADPH je potřebný na regeneraci glutathionu Oxidovaný glutathion vzniká při odbourávání peroxidu vodíku a organických peroxidů v erytrocytu. Hromadění peroxidů v buňce vyvolává hemolýzu 2GSH + HO-OH→ GS-SG + 2H2O GS-SG + NADPH + H+ 2GSH + NADP+ glutathionreduktasa Hromadění peroxidů v buňce vyvolává hemolýzu
Deficit glukosa-6-P dehydrogenasy v erytrocytech vrozená choroba způsobena bodovými mutacemi genu v chromosomu X u některých populací (až 400 různých mutací) až 400 milionů jedinců erytrocyty mají nedostatek redukovaného glutathionu choroba je často bezpříznaková a projeví se episodami hemolytické anemie po infekci, při užití některých léků (AAA)* nebo favových bobů (Vicia fava) Vysoká frekvence v tropických oblastech Afriky, Středomoří AAA* - antimalarika, antibiotika, antipyretika
Heinzova tělíska v erytrocytech s deficitem glukosa-6-P-dehydrogenasy Při deficitu redukovaného glutathionu dochází k poškození proteinů – částečná denaturace Vznikají nerozpustné proteiny vázané k membráně – Heinzova tělíska Erytrocyty jsou rigidní a nedeformovatelné – jsou odstraňovány makrofágy v játrech a slezině.
Favismus Hemolýza po požití bobů druhu Vicia fava
Metabolismus fruktosy Zdroj fruktosy: sacharosa z potravy, ovoce, med, HFCS Dříve příjem fruktosy 16-20 g/d, nyní se odhaduje až 85-100 g/d * HFCS (High fructose corn syrup). Vstup do buněk: GLUT V
Zásadní rozdíly mezi metabolismem glukosy a fruktosy • Fruktosa je metabolizována hlavně v játrech, enzymem fruktokinasou • Hexokinasa působí jen při vysoké koncentraci fruktosy • Játra metabolizují rychleji fruktosu než glukosu • Fruktosa sama nevyvolává uvolnění insulinu • Avšak příjem fruktosy může vyvolat následné zvýšení hladiny insulinu v důsledku přeměny fruktosy na glukosu
Metabolismus fruktosy 1 Převážná část fruktosy je metabolizována v játrech 2 Fruktosa ATP hexokinasa není regulovaná velmi nízké KM fruktokinasa Srovnejte s metabolismem glukosy Fruktosa- 1-P Fruktoso-6-P aldolasa B Zpětná konverze na glukosu Glyceraldehyd + dihydroxyaceton-P aldolasa B ATP triosa-kinasa glykolýza Glyceraldehyd-3-P
Aldolasa A a aldolasa B • jsou isoenzymy (je známa i aldolasa C) • aldolasa A : glykolýza (štěpení Fru-1,6-bisP) • aldolasa B: štěpení fruktosa-1-P • glukoneogenese (syntéza Fru-1,6-bisP)
1 Fruktokinasa a aldolasa B (játra): metabolismus obchází regulované enzymy glykolýzy (hexokinasu a fosfofruktokinasu) rychlé odbourání fruktosa je rychlý, na insulinu nezávislý zdroj energie • vysoký příjem fruktosy a její rychlý metabolismus na acetylCoA vede ke zvýšené tvorbě mastných kyselin a následně ke zvýšení produkce triacylglycerolů • vysoký příjem fruktosy může spotřebovat značné množství fosfátu na syntézu fruktosa-1-fosfátu. Fosfát chybí pro syntézu ATP.
Dříve se fruktosa doporučovala diabetikům jako neškodné sladidlo Současná doporučení: malé množství fruktosy v ovoci je neškodné slazení fruktosou nebo jiný vyšší příjem fruktosy (např. formou infuze) – není doporučeno
Poruchy metabolismu fruktosy Chybění aldolasy B – závažné !!! - hereditární fruktosová intolerance (autosomálně recesivní choroba) 1/40 000 Fruktosa-1-P se hromadí v buňkách jater → hypoglykemie Proč hypoglykemie? Hromadící se Fru-1-P inhibuje glykogenolýzu a glukoneogenezi Nutné včasné odhalení, objeví se po té, co kojenec začne přijímat sacharosu (v prvních týdnech života velmi závažné) (v některých zemích neonatální skrínink) Úprava dietou bez fruktosy a sacharosy
Poruchy metabolismu fruktosy Chybění fruktokinasy -benigní - esenciální fruktosurie Fruktosa se hromadí v krvi a vylučuje do moči Bez závažných projevů Dieta bez fruktosy Diagnostika: pozitivní redukční zkouška v moči průkaz glukosy specifickým testem negativní
Polyolová metabolická dráha - alternativní přeměna glukosy na glucitol a fruktosu v některých buňkách při vyšší koncentraci glukosy Glucitol nemůže z buňky unikat, pokud není přeměňován na fruktosu, hromadí se NADPH + H+ NADP+ D-glucitol D-glukosa aldosareduktasa NAD+ Játra, spermie, ovariální b. polyoldehydrogenasa NADH + H+ Chybí v čočce, retině, nervových buňkách Fruktosa (hlavní zdroj energie pro spermie)
Pozdní komplikace diabetu • při diabetické hyperglykemii vstupuje glukosa do buněk, které pro její vstup nevyžadují insulin • tvoří se D-glucitol, který nemůže z buňky unikat (past) • aktivita polyoldehydrogenasy v některých buňkách je malá (retina, čočka, nervová b.) • D-glucitol se hromadí • Zvýšený osmotický tlak vyvolává poruchy buněk (diabetická katarakta, retinopatie, neuropatie)
Metabolismus galaktosy zdroj: laktosa (mléko*) • přeměna v játrech na glukosu • epimerace • aby mohla proběhnout, je třeba galaktosu aktivovat: • na UDP-galaktosu * Ostatní mléčné výrobky (kefíry, tvaroh, sýry) laktosu téměř neobsahují
UDP-galaktosa (aktivní forma galaktosy) OH OH OH nevzniká přímou reakcí s UTP, nýbrž reakcí s UDP-glukosou
Metabolismus galaktosy v játrech Galaktosa je rychle metabolizována na glukosu Galaktosa ATP Galaktokinasa ADP UDP-glukosa Gal-1-P uridyltransferasa Glukosa-1-P UDP-galaktosa syntéza glykolipidů, GAG.. glykogen epimerasa UDP-glukosa
epimerasa UDP-galaktosa reakce je vratná, může být využita i k produkci glukosy Izomerace glukosy na galaktosu UDP-glukosa Význam galaktosy syntéza laktosy syntéza glykolipidů, proteoglykanů a glykoproteinů
Porucha metabolismu: „klasická“ galaktosemie • nedostatek uridyltransferasy - závažné !!!! • akumulace galaktosa-1-P a galaktosy • 1:40 000-60 000 živě narozených dětí, dědičnost onemocnění je autosomálně recesivní • interference s metabolismem fosfátů a glukosy, akumulace galaktosy a galaktosa-1-P v ledvinách, játrech, mozku a oční čočce • konverse galaktosy na galaktitol v oku – katarakta • nebezpečné pro novorozence, příznaky mezi 4.-9. dnem, hepatomegalie, ikterus, letargie, křeče • neléčená galaktosemie vede k poškození jater, ledvin, mozku • restrikce mléka a mléčných výrobků
Biosyntéza laktosy specifická pro mléčnou žlázu UDP-galaktosa Laktosa (galaktosyl-1,4-glukosa) glukosa Laktosasynthasa • Laktosasynthasa je komplex dvou proteinů: • galaktosyltransferasa (přítomná v mnoha tkáních) • -laktalbumin (přítomný v mléčné žláze jen při laktaci, syntéza je stimulována prolaktinem)
Metabolismus galaktosy v jiných buňkách Galaktosa a N-acetylgalaktosamin Jsou důležité komponenty glykoproteinů, proteoglykanů a glykolipidů. Syntéza těchto sacharidů probíhá ve všech typech buněk, galaktosyl andN-acetylgalaktosyl jsou přenášeny z UDP-galaktosy and UDP-N-acetyl-galaktosy pomocíUDP-galactosyltransferasy.
H O C H H O C H 2 P O C H 2 2 O O U T P O O H O H O H O O U D P H O O P H O O H H O H O H O H UDP-glukosa glukosa-1-P glukosa-6-P + N A D O O C H O 2 + N A D O O H glukosiduronáty O O U D P H O H UDP-glukuronát Metabolická dráha glukuronátu volný glukuronát glykosaminoglykany UDP-galakturonát UDP-iduronát
Příklady sloučenin vylučovaných ve formě glukosiduronátů Estrogeny Bilirubin Progesteron Meprobamat Morphin ad.
Odbourání D-glukuronové kyseliny Primáti a morčata NADPH + H+ NADP+ C O O H H O L-gulonát L-askorbát O H H O O H CO2 O H Kyselina D-glukuronová L-xylulosa blok: →esenciální pentosurie, pentosy v moči xylitol D-xylulosa může vstoupit do pentosového cyklu Xylulosa-5-P
C H O H 2 C H O H O O O O H H O H O H Syntéza L-askorbátu C H O H 2 1,4-lakton L-gulonové kyseliny C H O H - C O O O H O C H + H2O O 1 H H O C H H H C O H H O C H H H C L-gulonolaktonoxidáza -2H O H L-gulonát kys.askorbová
L-gulonolaktonoxidáza je neaktivní u primátů, a tedy i u člověka, ale též u ryb a ptáků. Aktivní u většiny obratlovců Je to výsledek ztrátové mutace (vlivem volných radikálů či virové infekce?) u předchůdce podřádu Anthropoidea asi před 25 milióny lety A co morče? Genom morčete obsahuje jiné pozůstatky genu pro gulonolaktonoxidázu. Fungovat tento gen přestal přibližně před 20 miliony let – morče tedy přišlo k své závislosti na vitaminu C jindy a jinak než my. Kyselinu askorbovou potřebují i rostliny.
Historie vitaminu C • 1754 – James Lind vydal knihu A Treatise on Scurvy, v níž navrhl citrony jako prevenci proti kurdějím. • 1761 – Východoindická společnost doporučila používat na svých lodích citrony a pomeranče. • 1795 – Bylo zavedeno preventivní podávání citronové šťávy na lodích anglického válečného námořnictva • 1911 – Kazimir Funk přišel s hypotézou, že příčinou některých závažných chorob může být nedostatek určitých nezbytných stopových látek – vitaminů –ve výživě. Prvními nemocemi, jejichž příčina byla hledána v avitaminóze, byly xeroftalmie (šeroslepost), beri-beri a kurděje; hypotetické vitaminy byly • označeny A, B a C. • 1928 – Albert von Szent-Györgyi izoloval kyselinu askorbovou • z papriky (Nobelovu cenu za to dostal r. 1937.) • 1933 – Tadeus Reichstein vyřešil chemickou syntézu kyseliny L-askorbové • 1957 – J. J. Burns objasnil biochemickou příčinu, proč člověk, opice a morče (na rozdíl od řady jiných obratlovců) nejsou schopni syntetizovat vitamin C. Dle VESMÍR 80, září 2001 497