Download

1 / 93
950 likes | 1.21k Views
+. ~ 30 Å. Ion Channels are the Valves of Cells Ion Channels are the Main Controllers of Biological Function. Ions in Water are the. Selectivity Different Ions carry Different Signals. Liquid of Life. Na +. Hard Spheres. Ca ++. Chemical Bonds are lines

E N D
+ ~30 Å Ion Channelsare theValves of CellsIon Channels are the Main Controllers of Biological Function Ions in Waterare the Selectivity Different Ions carry Different Signals Liquid of Life Na+ Hard Spheres Ca++ Chemical Bonds are lines Surface is Electrical Potential Redis negative (acid) Blueis positive (basic) K+ 3 Å 0.7 nm = Channel Diameter Figure of ompF porin by Raimund Dutzler
Ion Channelsare theValves of CellsIon Channels are the Main Controllers of Biological Function + ~30 Å Life occurs in ~130 mM salt solutions Selectivity Different Ions carry Different Signals Multiscale Na+ Hard Spheres Flow time scale is 0.1 msec to 1 min Averaging is in time, from 10-16 atomic scale to 10-4 biological scale! Averaging is in numberover 1012 water molecules needed to specify 10-7M concentrations and other ‘thermodynamic’ variables Ca++ K+ 0.7 nm = Channel Diameter 3 Å Chemical Bonds are lines Surface is Electrical Potential Redis negative (acid) Blueis positive (basic) Figure of ompF porin by Raimund Dutzler
Multiscale Issues more later Three Dimensional (106)3 Biological Scales Occur Together so must be Computed Together This may be impossible in simulations Physicists and Engineers have not tried
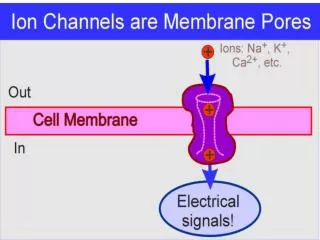
K+ ~30 Å Ion Channels are Biological Devices Natural nano-valves* for atomic control of biological function Ion channels coordinate contraction of cardiac muscle, allowing the heart to function as a pump Ion channels coordinate contraction in skeletal muscle Ion channels control all electrical activity in cellsIon channels produce signals of the nervous system Ion channels are involved in secretion and absorption in all cells:kidney, intestine, liver, adrenal glands, etc. Ion channels are involved in thousands of diseases and many drugs act on channels Ion channels are proteins whose genes (blueprints) can be manipulated by molecular genetics Ion channels have structures shown by x-ray crystallography in favorable cases *nearly pico-valves: diameter is 400 – 900 picometers
Thousands of Molecular Biologists Study Channels every day,One protein molecule at a timeThis number is not an exaggeration.We have sold >10,000 AxoPatch amplifiers Ion Channel Monthly AxoPatch 200B
Channels are parts of Machines, e.g., Excitation-Contraction Coupling L type Ca ChannelRyR ryanodine receptor L-type Ca Channel RyR Thanks for the figure toLászló Csernoch, Debrecen, Hungary Isabelle Marty, Grenoble, France
Single Channel Current open closed Slide from Mike Fill Thanks! Function of SINGLE isolated RyR Channels in Artificial Planar Lipid Bilayers AxoPatchPatch-Clamp Amplifier Designed at Rush Planar Bilayer Ca Fused Vesicle Experimental Chamber Teflon Septa 80-100 µM Diameter
Gating and Permeation Selectivity,Permeation are Amplitude Gating is Time Behavior
Channel Structure Does Not Change once the channel is open Amplitude vs. Duration Current vs. time Open Amplitude, pA 5 pA 100 ms Open Duration /ms Lowpass Filter = 1 kHz Sample Rate = 20 kHz Typical Raw Single Channel Records Ca2+ Release Channel of Inositol Trisphosphate Receptor : slide and data from Josefina Ramos-Franco. Thanks!
+ ~30 Å Channels are SelectiveDifferent Ions Carry Different Signals through Different Channels ompF porin Na+ Ca++ K+ 0.7 nm = Channel Diameter 3 Å Flow time scale is 0.1 msec to 1 min Figure of ompF porin by Raimund Dutzler
Channels are Selective Different Types of Channels use Different Types of Ionsfor Different Information
Central Problem* How does the channel control selectivity? *an example of “Reverse Engineering” For Modelers and Mathematicians:This is an inverse problem!
Goal: Understand Selectivity well enough toFit Large Amounts of Data*and to Make a Calcium Channel Atomic Scale Macro Scale *from many non-ideal solutions
Experiments have builtTwo Synthetic Calcium Channels MUTANT ─ Compound Calcium selective Unselective Wild Type As density of permanent charge increases, channel becomes calcium selectiveErev ECa built by Henk Miedema, Wim Meijberg of BioMade Corp.,Groningen, Netherlands Miedema et al, Biophys J 87: 3137–3147 (2004) Mutants of ompF Porin Designed by Theory Glutathione derivatives Atomic Scale Macro Scale
Central Problem* How does the channel control selectivity? “Reverse Engineering” This is an inverse problem Closely related inverse problemshave been solved by mathematics used to design blast furnaces Burger, Eisenberg, and Engl (2007) SIAM J Applied Mathematics 67:960-989
Selective Binding CurveL type Ca channel Wolfgang Nonner
For Modelers and Mathematicians:This is reverse engineering! Inverse Problem for Selectivity Badly posed, simultaneously over and under determined with noise and systematic error has actually been solved using methods for the Inverse Problem of a Blast Furnace Burger, Eisenberg and Engl (2007) SIAM J Applied Math 67: 960-989
Channels are only Holes Why can’t we understand and build them? Helpful to know physical basis of functionif we want to build one and improve it Where do we start? Not with Molecular Mythology Not with gas phase models of traditional channology Liquids are not Gases; biological solutions are not ideal Not with guesses about trajectories of structural biologistsCounting, Statistics, and Averaging are Essential
Why can’t we understand and build channels? Uncalibrated Simulations will not make devices that actually work Unpopular view because Calibration is Hard Work
Multiscale Issuesare the key if we want to actually build channels that work Three Dimensional (106)3
Multiscale Issues Three Dimensional (106)3 Biological Scales Occur Together so scales must be CALIBRATED TOGETHER in real biological solutions that are MIXTURES OF IONS
Multiscale Issues Calibrations of Molecular Dynamics in Real Solutions are just starting! Unpopular Reality: hard work It may not be possible to deal with scale ratios of 1011 , 106 ,1018 , 1012 all at once
Multiscale Issues It may not be possible to deal with Ratios of Scales of 1011 106 1018 1012 all at once Physicists and Engineers do not even try!
Channels are only Holes Why can’t we understand and build them? Where do we start? Science as Usual Guess and Check
Working Hypothesis Biological Adaptation is Crowded Ions and Side Chains
Active Sites of Proteins are Very Charged 7 charges ~ 20M net charge = 1.2×1022 cm-3 Pure water is 55 M + + + + + - - - - Selectivity Filters and Gates of Ion Channels are Active Sites Physical basis of function OmpF Porin Hard Spheres Na+ Ions are Crowded K+ Ca2+ Na+ Induced Fit of Side Chains K+ 4 Å Figure adapted from Tilman Schirmer
Working Hypothesis Biological Adaptation is Crowded Ions and Side Chains Everything interactsClassical Modelsand Force Fields of Molecular Dynamics assume no interactions with ion concentrations
Finite Size EffectsWorking Hypothesis Chemically Specific Properties of ions (e.g. activity = free energy per mole) come from interactions of their Diameter and Charge and dielectric ‘constant’ of ionic solution Atomic Detail ‘Primitive Implicit Solvent Model’ learned from Doug Henderson, J.-P. Hansen, Stuart Rice, among others…Thanks!
Ions in Water are the Liquid of Life. They are not ideal solutions Chemically Specific PropertiesofIonic Solutions come fromInteractions Molecular Dynamics Force Fields are Calibrated assuming no interactions with concentrations Force Fields must be REcalibratedin each Biological Solution
Ions in Water are the Liquid of Life. They are not ideal solutions Chemically Specific PropertiesofIonic Solutions come fromInteractions Chun Liu’s Energetic Variational Principle deals with Interactions EnVarA
Ions in Water are the Liquid of Life They are not ideal solutions Everything Interacts with Everything For Modelers and Mathematicians Tremendous Opportunity for Applied MathematicsChun Liu’s Energetic Variational Principle EnVarA
Ions in Water not ideal solutions Chemically Specific Properties Come from Interactions Chun Liu’s Energetic Variational Principle deals with Interactions EnVarA Variational Principles Deal with Multiple Scales and Interactions Consistently and Automatically
Variational Principles Deal with Multiple Scales Consistently and Automatically EnVarA New Component or Scaleof Energy or Dissipation implies New Field Equations (Euler Lagrange) by Algebra Alone No new Assumptions
Energetic Variational ApproachEnVarAChun Liu, Yunkyong Hyon, and Bob Eisenberg Mathematicians and Modelers: two different ‘partial’ variations written in one framework, using a ‘pullback’ of the action integral CompositeVariational Principle Action Integral, after pullback Rayleigh Dissipation Function Euler Lagrange Equations Field Theory of Ionic Solutions that allows boundary conditions and flow and deals with Interactions of Components self-consistently
Energetic Variational AnalysisEnVarAbeing developed by Chun Liu with creates a newMultiscaleField Theory of Interacting Components that allows boundary conditions and flow and deals with Ions in solutions self-consistently (1) Yunkyong Hyon, Bob Eisenberg. Ions in Channels (2) Rolf Ryham, Bob Eisenberg and Fred Cohen. Virus fusion toCells (3) Yoichiro Mori and Bob Eisenberg. Water flow in Tissues Multiple Scales
We have already established that the Implicit Solvent (“Primitive”) Model ofIonic Solutions Describes Calcium and Sodium Channels quite well at EquilibriumwithoutPreformed Structure Structure is the Computed Consequence of the Model
Selectivity FilterCrowded with Charge Selectivity Filter O½ Wolfgang Nonner L type Ca Channel + ++ “Side Chains”
Multiscale Analysis at Equilibrium Solved with Metropolis Monte Carlo MMC Simulates Location of Ionsboth the mean and the variance ProducesEquilibrium Distribution of location of Ions and ‘Side Chains’ MMC yields Boltzmann Distribution with correct Energy, Entropy and Free Energy Other methodsgive nearly identical results: Equilibrium MultiscaleMSA (mean spherical approximation SPM (primitive solvent model) DFT (density functional theory of fluids), Non-equilibrium Multiscale DFT-PNP (Poisson Nernst Planck) EnVarA…. (Energy Variational Approach) etc
Metropolis Monte Carlo Simulates Location of Ionsboth the mean and the variance Details: • Start with Configuration A, with computed energy EA • Move an ion to location B, with computed energy EB • If spheres overlap, EB → ∞ and configuration is rejected • If spheres do not overlap, EB→ 0 and configuration is accepted • If EB < EA: accept new configuration. • If EB > EA : accept new configuration with probability Key idea MMC chooses configurations with a Boltzmann probability and weights them evenly instead of choosing them from uniform distribution and then weighting them with exp(−E/k BT)
Crowded Ions Snap Shots of Contents Radial Crowding is Severe ‘Side Chains’are SpheresFree to move inside channel 6Å Parameters are Fixed in all calculations in all solutions for all mutants Experiments and Calculations done at pH 8 Boda, Nonner, Valisko, Henderson, Eisenberg & Gillespie
Mutation • Na Channel • Ca Channel Same Parameters • E • E • E • A • D • E • K • A Charge -3e Charge -1e 1 0.004 Na+ Ca2+ Na+ Occupancy (number) 0.5 0.002 Ca2+ 0 0 -6 -4 -2 0 0.05 0.1 log (Concentration/M) Concentration/M EEEE has full biological selectivityin similar simulations Boda, et al
Calcium Channelhas been examined in ~35 papers, e.g., Most of the papers are available at ftp://ftp.rush.edu/users/molebio/Bob_Eisenberg/Reprints http://www.phys.rush.edu/RSEisenberg/physioeis.html
Calcium Channel Summary Simulation Paper (and target for new experiments!) in Experimental Journal Next, the Sodium Channel
Next, the Sodium Channel specifically, the Aspartate DAcid Negative Glutamate EAcid Negative Lysine KBasicPositive Alanine A Aliphatic Neutral DEKASodium Channel 6 Å
DEKASodium Channel has very different properties from Ca channel,e.g., ‘binding’ curve,Na+ vs Ca++ selectivity Na+ vs K+ selectivity
Challenge from leading biophysicists Walter Stühmer and Stefan Heinemann Max Planck Institutes, Göttingen, Leipzig Can a physical theory explain the mutation DEEA into DEKA?
Mutation • Na Channel • Ca Channel Same Parameters • E • E • E • A • D • E • K • A Charge -3e Charge -1e 1 0.004 Na+ Ca2+ Na+ Mutation Occupancy (number) 0.5 0.002 Same Parameters Ca2+ 0 0 -6 -4 -2 0 0.05 0.1 log (Concentration/M) Concentration/M EEEE has full biological selectivityin similar simulations Boda, et al
Nothing was changedfrom the EEEA Ca channelexcept the amino acids Calculated DEKA Na Channel SelectsCa 2+vs.Na + and also Na+vs. K+ Calculations and experiments done at pH 8