Download
1 / 12
120 likes | 243 Views
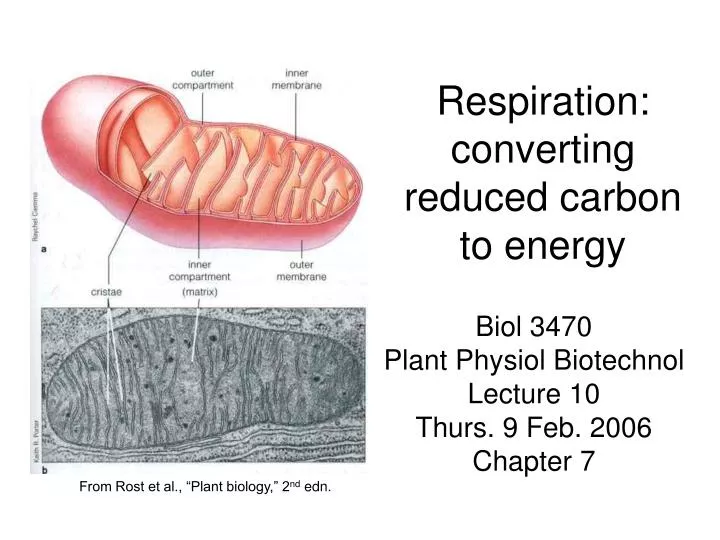
Respiration: converting reduced carbon to energy. Biol 3470 Plant Physiol Biotechnol Lecture 10 Thurs. 9 Feb. 2006 Chapter 7. From Rost et al., “Plant biology,” 2 nd edn. Cellular respiration oxidizes fixed carbon to CO 2.

E N D
Respiration: converting reduced carbon to energy Biol 3470 Plant Physiol Biotechnol Lecture 10 Thurs. 9 Feb. 2006 Chapter 7 From Rost et al., “Plant biology,” 2nd edn.
Cellular respiration oxidizes fixed carbon to CO2 • Photosynthesis has fixed (reduced) atmospheric carbon into carbohydrates (photoassimilate) • Respiration mobilizes the energy in these macromolecules using oxidative enzyme reactions via • Glycolysis (6C 3C) • The citric acid cycle or CAC (a/k/a the TCA cycle) (using the 3C molecules to generate reducing power [electrons] for…) • Oxidative phosphorylation: making ATP • Photoassimilate also represents a source of C skeletons for anabolism • When we discuss respiration, we will concentrate on plant respiration and cover: • Interesting differences between plant and animal respiration • Influence of environmental factors on respiration • The relationship of respiration to plant productivity (yield)
Compare and contrast respiration and photosynthesis • Identical and reciprocal substrates and products but • Occur in different cellular compartments • Use different enzymes • The two processes are extensively linked Dependent on photosynthesis! Note that plastids have a parallel set of carbohydrate metabolizing enzymes! CAC + oxidative P’n (respiration) Gluco- neogen-esis Glyco- lysis Gluco- neogenesis Glyco- lysis Fig. 7.1
To fuel respiration, starch must be moved into the cytosol Favored energetically! • In most species, starch is the main “fuel” for respiration • Starch is degraded where it is synthesized: in __________ • But glycolysis takes place (mostly) in cytosol • need to export starch hydrolysis products into cytosol This occurs by 2 possible routes: • As Glucose by a glucose transporter • As triose-P by the triose-P/Pi antiporter already discussed • Sucrose breakdown in cytosol occurs via • Sucrose synthase • Recall that the enzyme’s high DG for sucrose synthesis actually favors its breakdown (+14kJ/mol) • Invertase: alkaline (cytosol), acid (cell walls, vacuoles) Starch phosphorylase amylase Glc-1-P Plastidic glycolysis 1 2 1 Triose -P trans- porter glucose transporter 2 Cytosolic Figure 7.3
The net products of the oxidation of hexoses are water, CO2 and ATP • We will not rehash the basics of respiration in any detail • See section 7.1 for summary, sections 7.3 to 7.7 to review in detail • Know that entropy greatly favors hexose oxidation over its synthesis • Plants may get from hexose to CO2 the same way (using the same enzymes) as animals • However, plants need to be more metabolically flexible than animals because of their sessile nature • Adaptation to their environment means coping with abiotic (“non-living”) stresses • In plants, this often means using different enzymes to accomplish the same metabolic goals easily performed in homeothermic animals
Plants possess “bypass” enzymes for regulatory steps of glycolysis 1 • The activity of regulatory enzymes in glycolysis is often limited under nutrient stress • e.g., Pi stress: occurs often in nature! • Complete plant fertilizer is “NPK” • These “bypass” or alternate enzymes allow glycolysis to progress even in the absence of Pi Examples: • PFP to bypass PFK • nonphosphorylating NADP-G3PDH to bypass Pi requiring equilibrium enzymes • PEP carboxylase as part of a bypass of pyruvate kinase 2 3 1 2 3 Theodorou and Plaxton 1993
The activity of alternative oxidase generates heat instead of ATP Normally donates its e- to complexes II III IV • The terminal electron acceptor in mitochondrial electron transport is normally c_______ C oxidase (a/k/a Complex IV) • Its activity is inhibited by CN ( ______ ), CO, N3- in animals and plants Figure 7.12 • These are effective poisons in animals • However, plants possess a CN- resistant oxidase called the alternative oxidase • Accepts electrons from UQ pool (bypasses complexes II-IV) and transfers electrons directly to oxygen • Bypasses stepwise electron reduction of energy allowing H+ extrusion and ATP synthesis!
The activity of the alternative oxidase (AOX) may be used to generate heat • The activity of the AOX severely limits ATP synthesis • only allows 2 H+ to exit the mitrochondria’s matrix to generate 0 or 1 ATP • Compared to ~9 H+ 3 ATP in oxidative phosphorylation So why use it? • Reason 1: It may raise the temperature of plant tissues up to 10°C above ambient • This thermogenesis volatilizes amines that attract pollinators (skunk cabbage and other fly-pollinated species of the Arum family) • Reason 2: Energy overflow hypothesis AOX engages only after photosynthesis derived substrates saturate the oxidative E.T.C. • This allows the plant to • burn off excess fixed C that interferes with source-sink relationships and inhibits photoassimilate translocation • prevent overreduction (lots of e-) of mt E.T.C. which causes production of ROS (e.g. superoxide) Skunk cabbage. From Rost et al., “Plant biology,” 2nd edn.
Oilseeds are able to convert stored oil to carbohydrate • Many seeds store a significant portion of photoassimilate as oil, not carbohydrate • This oil is mobilized as an energy source upon germination • e.g., canola (45% oil by dry weight versus maize 5%) cbc.ca • Oil – not water soluble, not transportable • Most plants convert oil droplets (triglycerides) sucrose to mobilize its energy • Animals cannot interconvert lipids and carbohydrates! • Again, this gives plants metabolic flexibility in allocating carbon between lipids and carbohydrates • Seeds can be smaller because lipids store more energy per gram!
Mobilizing the energy in stored oil involves the glyoxylate cycle and gluconeogenesis Figure 7.13 • Triglyceride conversion to sucrose involves 3 organelles + cytosol • Fatty acids are removed from triglyceride by lipase • FA imported into glyoxysome – specialized plant organelle • Cleaved at every 2nd C to generate acetyl CoA via ß-oxidation • Glyoxylate cycle take home messages: • Borrowing oxaloacetate from the mitrochondrion allows citrate synthesis from fatty acids • It’s a cycle! Regeneration of OAA in mt keeps acetyl CoA incorporation high • The products of the cycle enter gluconeogenesis to generate sucrose in the __________ • Glycerol from triglyceride also enters gluconeogenesis for sucrose biosynthesis • NADH enters oxidative phosphorylation a/k/a _____________
Respiration rate varies depending on the plant tissue Figure 7.15 • Respiration rate varies with plant age • Related to metabolic demand • High during active growth • Falls during maturation of tissues • Difficult to relate to individual pathway flux in mature plants – varies with nutrient status, organ maturation state, temperature • Senescing tissues increase CO2 evolution (e.g., in many ripening fruits) • This is known as the respiratory climacteric • Accompanied by “uncoupling” of oxidative phosphorylation from ATP synthesis • Respiration accounts for 30-60% of photoassimilate lost as CO2 30-70% of this is from roots! http://tfphotos.ifas.ufl.edu/022401.HTM
Environmental effects also change the respiration rate • Light • Rate in light not always = dark rate • Difficult to measure respiration in light during photosynthesis due to O2 evolution, CO2 recycling • higher in shade-intolerant vs tolerant species (low irradiance low respiration) • Due to/cause of lower growth rate in shade-grown plants? • Temperature • Respiration rises exponentially from 5 30°C, doubles with every 10°C rise • This parameter can be defined as the temperature coefficient Q10(=2) • Optimum temperature for respiration varies depending on environment • 10°C for arctic species, 30°C for tropical species • Likely due to other factors limiting respiration (ATP availability, different temperature optima for photosynthetic enzymes) • Oxygen • Cytochrome C (terminal electron acceptor) has high affinity for O2 • Normally its high concentration is not limiting except in bulky tissues (fruit, roots) – air spaces circulate gases, flooding causes anoxia