Download

1 / 34
350 likes | 572 Views
Bacterial Protein Translocation & Pathogenesis. David R. Sherman HSB G-153 221-5381 dsherman@u.washington.edu. Lecture outline. Cellular addresses Getting stuck in the membrane: YidC Crossing the inner membrane: Sec-dependent SRP Sec B TAT-mediated

E N D
Bacterial Protein Translocation & Pathogenesis David R. Sherman HSB G-153 221-5381 dsherman@u.washington.edu
Lecture outline • Cellular addresses • Getting stuck in the membrane: YidC • Crossing the inner membrane: • Sec-dependent • SRP • Sec B • TAT-mediated • Crossing the outer barrier - specialized secretion systems • An example in gram (+) bacteria (paper discussion)
Protein destinations Approx 10% of proteins cross at least the inner membrane. Approx 30% of proteins are membrane associated.
Machinery of bacterial protein translocation Cytosolic membrane (Gram +/-): YidC. Sec machinery. Tat translocation. Cell wall (Gram +/-): very little known. Outer membrane (Gram -): several specialized systems. Much better studied in Gram-negatives.
Membrane insertion via YidC • Multi-pass membrane protein. • Needed for insertion of some (all?) membrane proteins. • Can act alone or w/ Sec YEG. • Signal sequence at N-terminus. • Evolutionary origin of secretion?
Across the cytoplasmic membrane -- the Sec machinery General features: Sec YEG: heterotrimeric pore-forming membrane proteins. SecA: membrane-associated ATPase. Substrates are generally unfolded. Substrates have a signal peptide: usually N-terminal 1(+) basic AAs followed by10-20 hydrophobic AAs
SecYEG topology 9:494-500, 2001 Homologous to eukaryotic Sec61p complex.
SRP-mediated translocation SRP: homologous to eukaryotic SRP Ffh (54 homolog) 48kDa GTPase ffs (4.5S RNA) essential for cell viability Recognizes ribosome-bound nascent membrane proteins. Substrate recognition is via signal sequence. SecA is NOT needed for membrane association, but IS needed for translocation.
SecB-mediated translocation SecB: acidic, cytosolic chaperone. recognizes “mature”, unfolded proteins. destination -- periplasm, outer membrane or beyond. Substrate recognition is NOT via the signal sequence. Binding motif: ~9 AAs long. hydrophobic and basic. acidic AAs strongly disfavored.
SecB protein targeting Nat Struct Biol. 2001 8(6):492-8.
Sec interactions 9:494-500, 2001
Twin-arginine (Tat)-mediated protein translocation Independent of the Sec system. TatA and TatC are essential. Transports folded proteins. Not found in eukaryotes; some bacteria. # of Tat substrates per organism varies very widely -- None (Clostridium tetani, Fusobacterium) 145 (Streptomyces coelicolour)
Twin-arginine (Tat)-mediated protein translocation Targeting signal (in the first 35 AAs) has 3 regions: N-term is positively charged (S/T)-R-R-x-F-L-K hydrophobic a-helical domain C (cleavage) domain. TATFIND 1.2
Inner membrane hierarchy • Co-translational: • SecYEG; Sec A, SRP • Very hydrophobic N-terminal signal • Newly synthesized, unfolded: • SecYEG, SecA, SecB • Hydrophobic N-terminal signal • Fully folded: • Tat, Hydrophobic N-terminal signal w/ RR • Very little cross-talk!
Specialized secretion pathways (Gram-negative bacteria) 1. Type I pili. 2. Type I secretion. 3. Type II secretion/general secretory pathway/type IV pili. 4. Type III secretion (TTSS). 5. Type IV secretion. 6. Type V/autotransporters. Classification is based on the sequence/structure of the transport machinery and their catalyzed reactions. These systems are usually associated with virulence.
Assembly of type I pili Uropathogenic E. coli Allow for attachment during the initial stages of infection. Assembled in two stages: Sec-dependent Pap C/D-dependent
Type I secretion of repeat toxins: E. coli HlyA Lipid-modified toxin. 11-17 repeats of 9 AAs. Binds Ca++ Punches holes. Sec-independent. Requires ABC-transporter (HlyB). C-term signal sequence.
Type II secretion -- the “general” secretory pathway Many examples: **cholera toxin** alkaline phosphatase proteases elastase Type IV pili Occurs in 2 steps -- 1st is Sec-dependent; 2nd requires 10 proteins and ATP. Secretion signal?
Type III secretion Needle Triggered by contact w/ host cells. Sec-independent, similar to flagellar assembly. Assembly of the needle occurs at the tip.
The type III needle complex Nature 435, 702-707 (2 June 2005)
Type IV secretion Pertussis toxin Agarobacterium Very versatile; Sec- and ATP-dependent.
Autotransporters: Neisseria IgA protease Also called type V secretion Synthesized as a pre-proenzyme. C-term b-barrel inserts in OM, pulls N-term through. N-term auto-cleaves, promoting release.
Cell wall proteins of Gram (+)s Initially Sec-dependent. N-term signal cleaved. C-term signal sorts to CW. L-P-x-T-G. Amide linkage to peptidoglycan.
So what’s in common? • All secretion systems must: • assemble themselves. • recognize the appropriate substrates. • maintain proper folding state. • determine their final locations.
Additional reading (not assigned) The structural basis of protein targeting and translocation in bacteria. Driessen AJ, Manting EH, van der Does C. Nat Struct Biol. 2001 8(6):492-8. The Tat protein export pathway. Berks BC, Sargent F, Palmer T. Mol Microbiol. 2000 Jan;35(2):260-74. Prokaryotic utilization of the twin-arginine translocation pathway: a genomic survey. Dilks K, Rose RW, Hartmann E, Pohlschroder M. J Bacteriol. 2003 Feb;185(4):1478-83. Protein secretion and the pathogenesis of bacterial infections. Lee VT, Schneewind O. Genes Dev. 2001 Jul 15;15(14):1725-52. Getting out: protein traffic in prokaryotes. Pugsley AP, Francetic O, Driessen AJ, de Lorenzo V. Mol Microbiol. 2004 Apr;52(1):3-11.
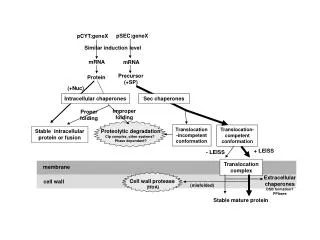
Fig 1. Guinn et al, Mol Microbiol, 2004, 51(2):359-370.
Fig. 2 Guinn et al, Mol Microbiol, 2004, 51(2):359-370.
Fig. 3 Guinn et al, Mol Microbiol, 2004, 51(2):359-370.
Fig. 4 Guinn et al, Mol Microbiol, 2004, 51(2):359-370.
Fig. 5 Guinn et al, Mol Microbiol, 2004, 51(2):359-370.
Fig. 6 Guinn et al, Mol Microbiol, 2004, 51(2):359-370.
Fig. 7 Guinn et al, Mol Microbiol, 2004, 51(2):359-370.