Download

1 / 28
320 likes | 1.06k Views
Genetic drift. Guest lecture by Kim van der Linde. Hardy-Weinberg equilibrium. Infinite populations Gene frequencies remains constant over generations What if populations are finite? Sampling error. Genetic drift versus natural selection. Selection: fitness of alleles differs

E N D
Geneticdrift Guest lecture by Kim van der Linde
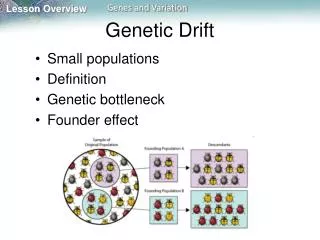
Hardy-Weinberg equilibrium • Infinite populations • Gene frequencies remains constant over generations • What if populations are finite? • Sampling error
Genetic drift versusnatural selection • Selection: fitness of alleles differs • Drift: fitness of alleles is equal => (Selectively) Neutral
Genetic drift • Random change in the allelic frequencies that results from the sampling of gametes from generation to generation
Evolutionary important? • Bottlenecks • Northern elephant seal (no variation in enzyme-coding loci => 20 individuals in 1890, now 30,000) • Founder effects • Island species
Two approaches • Random walk • Coalescence
Random walk: Model 1 • Wright-Fisher model • Non-overlapping generations • Binomial distribution • Selfing allowed (makes the statistics easier) • Each parent has equal chance • No selection, gene-flow or mutation • Named after Sewall Wright and R.A. Fisher
Random walk : Model 2 • Moran model • Overlapping generations • Short time intervals: • one individual reproduces (+ 1 allele) • one individual dies (- 1 allele) • can be the same individual; net no change • No selection, gene-flow or mutation
Binomial distribution Flip a coin • p = .5 • q = 1-p = .5
Binomial distribution • j = # A2alleles in generation t • i = # A2alleles in generation t+1 • N = population size
2/32 16/32 8/32
Fixation across populations • What is the probability of fixation for each allele? • Many populations => infinite • Hardy-Weinberg equilibrium • No change in frequency • So, probability of fixation is equal to initial frequency • Formally, Markov transition probability matrix
Theoretical expectation for drift over many populations Hartl and Clark. 1989. Principles of Population Genetics. Sinauer, Sunderland MA.
Coalescence • Derivations of the gene copies in one or more populations from a single ancestral copy, viewed retrospectively
Coalescence: family names Grandparents Uncle Aunt Father Me Brother Cousin ♂ Daughter
No drift in a perfect world, where everyone has the same number of offspring. We don’t live in a perfect world.
reference population most recent common ancestor
Identity by state <->Identity by decent allozygous autozygous
Co-ancestry • f: the coancestry coefficient or coefficient of inbreeding = probability that two genes are ibd • F: departure from HW • With random mating: f = F • H=1-F • Generation 0: F0 = 0, H0 = 1
Co-ancestry Chance of drawing same allele twice (n = 2):
Co-ancestry • f: the coancestry coefficient or coefficient of inbreeding = probability that two genes are ibd • F: departure from HW • With random mating: f = F • H=1-F • Generation 0: F0 = 0, H0 = 1 • Generation 1: F1 = 1/2N; H1 = 1- 1/2N
T=4N (diploid) or T=2N (haploid) Expected time to a common ancestor in generations is twice the number of alleles