Download

1 / 0
10 likes | 140 Views
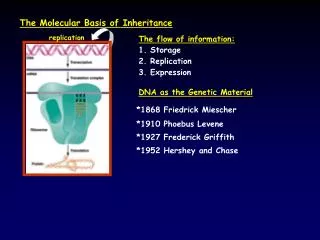
The Molecular Basis of Inheritance. Campbell and Reece Chapter 16. The Search for Genetic Material. once Morgan proved genes are in chromosomes big debate started: Is the genetic material in chromosomes the DNA or the proteins? @ first case for proteins seemed stronger very heterogenous

E N D