Download
1 / 45
450 likes | 580 Views
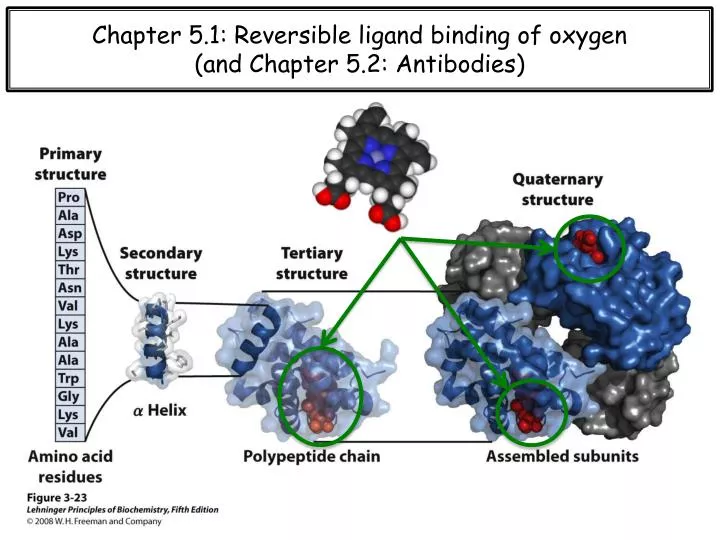
Chapter 5.1: Reversible ligand binding of oxygen (and Chapter 5.2: Antibodies). When twenty just isn ’ t enough….

E N D
Chapter 5.1: Reversible ligand binding of oxygen (and Chapter 5.2: Antibodies)
When twenty just isn’t enough… (Modified) From Wikipedia: A cofactor is a non-protein molecule that is bound to a protein and is required for the protein's biological activity. Sometimes with enzymes, the term coenzyme is used instead. Note that some sources limit the use of the term "cofactor" to inorganic substances, but I do not. A protein without its cofactor is called an apo-protein, whereas the complete protein + cofactor complex is the holo-protein. AMP Some enzymes or enzyme complexes require several cofactors. For example, the multi-subunit pyruvate dehydrogenase complex requires 5 organic cofactors and 1 metal ion. Organic cofactors are often vitamins or are made from vitamins. Many include the nucleotide AMP scaffolding, which may reflect a common evolutionary origin as part of ribozymes in the ancient world.
Heme is a prosthetic group that consists of an iron atom contained in the center of a heterocyclic porphyrin molecule
A primer on chemical kinetics [A] [P]
Quantifying reversible ligand binding with the association equilibrium constant
The globin fold serves as a scaffolding for heme Myoglobin a-Hemoglobin b-Hemoglobin
Based on common evolutionary descent and similar function, globin sequences are quite similar
Heme serves as a scaffolding for iron, which binds oxygen Distal His Proximal His
The roles of the distal histidine CO binds to free heme many, many orders of magnitude stronger than O2, which would result in suffocation simply due to ambient levels of CO in the atmosphere. vs. However, in globins the affinity of CO for heme is only ~ 250x stronger than O2. Secondarily, the distal His prevents oxidation of the iron by any number of possible cellular oxidizing agents. Distal HIS Distal HIS Note that the Fe-CO interaction is not what you might expect. Due to a phenomenon known as p-backbonding, the carbon of CO actually ligates iron.
Hemoglobin (Hb) vs. myoglobin (Mb) q Note that concentrations of gases are normally expressed as partial pressures. pO2 (kPa)
T vs. R hemoglobin Oxygen reservoir
Oxygen binding initiates a conformational change that is propagated through the Hb interaction network
Quantifying cooperativity with the Hill coefficient The Hill plot should have a slope of n. However, the experimentally determined slope doesn’t actually reflect the number of binding sites. Rather, nH is the interaction between the binding sites, which is a measure of cooperativity.
Cooperativity is a subset of allostery, which is a generic term indicating long-range intramolecular communication From Wikepdia: In biochemistry, allosteric regulation is the regulation of an enzyme or other protein by binding an effector molecule at the protein's allosteric site (that is, a site other than the protein's active site). Effectors that enhance the protein's activity are referred to as allosteric activators, whereas those that decrease the protein's activity are called allosteric inhibitors. The term allostery comes from the Greek allos, "other," and stereos, "solid (object)," in reference to the fact that the regulatory site of an allosteric protein is physically distinct from its active site. Allosteric regulations are natural example of control loops, such as feedback from downstream products or feedforward from upstream substrates.
How does allostery occur? Q: How do allosteric signals get propagated over distances of tens of Å? ? A simple and common description, which may or may not be correct.
MWC vs. KNF models of Hb cooperativity (all or none vs. sequential)
How can we affect Hb-O2 binding to access the ‘reservoir’? Hb + 4 O2 Hb.(O2)4 vs. Hb + 4 O2 Hb.(O2)4 Oxygen reservoir
Method #1: 2,3-bisphosphoglycerate BPG causes an alteration in the structure of HB, which, causes it to give up its O2. Another way of saying this is that the Hb-O2 dissociation curve 'shifts to the right’ (as shown). This means that at a given partial pressure of oxygen, the percent saturation for Hb with be lower.
Method #1: 2,3-bisphosphoglycerate • Binding of BPG to deoxyHb: • BPG binding stabilizes the T state of deoxyHB, shown here as a mesh surface image. • The negative charges of BPG interact with several positively charged groups (shown in blue in this surface contour image) that surround the pocket between the β subunits in the T state. • (c) The binding pocket for BPG disappears on oxygenation, following transition to the R state.
Method #2: The Bohr effect (Hb releases O2 with increasing [H+]) When a skeletal muscle starts contracting, the cells in that muscle use more oxygen, make more ATP, & produce more waste products (CO2). More CO2 translates into a lower pH. That is so because this reaction occurs when CO2 is released: CO2 + H20 → H2CO3 → HCO3- + H+
Method #2: The Bohr effect (Hb releases O2 with increasing [H+])
Sickle cell anemia There are large number of genetic variations in the Hb subunits, all of which affect oxygen binding. Sickle cell anemia: HbS is variant of b-globin, caused by a single nonsynonymous mutation within the ORF (E6V). SCA is the most common inherited blood disorder in the United States, affecting about 72,000 Americans or 1 in 500 African Americans. SCA is characterized by episodes of pain, chronic hemolytic anemia and severe infections, usually beginning in early childhood. The abnormal HbS clusters together, distorting the RBCs into sickled shapes. These deformed and rigid RBCs become trapped within small blood vessels and block them, producing extreme pain and eventually damaging organs.
The sickle cell mutation causes a structural change within Hb
Hb-S aggregates in order to ‘hide’ its hydrophobic surface valine from solvent
Sickle cell confers an evolutionary resistance to malaria The precise mechanism by which sickle cell trait imparts resistance to malaria is unknown. A number of factors likely are involved and contribute in varying degrees to the defense against malaria. Red blood cells from people heterozygous with sickle trait do not sickle to any significant degree at normal venous oxygen tension. Very low oxygen tensions will cause the cells to sickle, however.
Sickle cell confers an evolutionary resistance to malaria • As shown in the life cycle of Plasmodium in the figure (previous page), an asexual stage of the organism lives in red blood cells in humans, while a sexual phase develops in the mosquito. • The asexually-reproducing forms, or merozoites, develop within red blood cells, and then break out to infect new cells. • While reproducing asexually inside the red blood cells, the merozoites have a high metabolic rate and consequently consume lots of oxygen. • If the individual is heterozygous for sickle-cell trait, half their hemoglobin is HbA-HbS. Thus it will sickle when the oxygen tension becomes very low inside the red blood cells (sickling does occur in heterozygous individuals, but at a lower oxygen tension than for homozygotes). • These sickled cells are removed from the body by the spleen, along with the merozoites inside of them. • Thus heterozygotes on the average remove merozoites from their body before the microorganisms have a chance to produce a large infectious population inside the body. • It is this selective advantage of the heterozygote that maintains the HbS gene at a higher level in malarial than in non-malarial environments.
Ch. 5.2: Antibodies • From Wikipedia: • An antibody also known as an immunoglobulin is a large Y-shaped protein used by the immune system to identify and neutralize foreign objects like bacteria and viruses. • The antibody recognizes a unique part of the foreign target, termed an antigen. • Each tip of the "Y" of an antibody contains a paratope (a structure analogous to a lock) that is specific for one particular epitope (that is equivalent to a key) on an antigen, allowing these two structures to bind together with precision. • Using this binding mechanism, an antibody can tag a microbe or an infected cell for attack by other parts of the immune system, or can neutralize its target directly (for example, by blocking a part of a microbe that is essential for its invasion and survival).
Antibodies are constructed from immunoglobin (Ig) domains From Wikipedia: The immunoglobulindomain is a type of protein domain that consists of a 2-layer sandwich of between 7 and 9 antiparallel β-strands arranged in two β-sheets with a Greek key topology. The backbone switches repeatedly between the two β-sheets. Members of the immunoglobulin superfamily are found in hundreds of proteins of different functions. Examples include antibodies, the giant muscle kinase titin and CuZnSOD.
Antibody-antigen interactions Antibody-antigen complexes are stabilized by the same types of interactions that stabilize protein structure and other protein-substrate complexes.
Antibody-hapten interactions From Wikipedia: A haptenis a small molecule that can elicit an immune response only when attached to a large carrier such as a protein. The carrier may be one that also does not elicit an immune response by itself. Note that, in general, only large molecules, infectious agents, or insoluble foreign matter can elicit an immune response in the body.