Download

1 / 47
470 likes | 586 Views
The role of aquaporins in the freeze tolerance of yeast cells: application in frozen dough. Patrick Van Dijck. Department of Molecular Microbiology VIB Laboratory of Molecular Cell Biology K.U.Leuven. Excellent University, Bratislava 6 May 2008. VIB: Mission & Objectives. mission

E N D
The role of aquaporins in the freeze tolerance of yeast cells: application in frozen dough Patrick Van Dijck Department of Molecular Microbiology VIB Laboratory of Molecular Cell Biology K.U.Leuven Excellent University, Bratislava 6 May 2008
VIB: Mission & Objectives • mission • to conduct world class biomolecular research for the benefit of scientific progress and society • objectives • strategic basic research • translate research results into products • public outreach & education
Antwerp Ghent Brussels Louvain VIB’s Road to Success • 1000 scientists and technicians • 65 research groups in 8 departments
Red biotechnology Green biotechnology • intestinal glucose sensing • antifungal targets • human diseases • trehalose metabolism • sugar sensing White biotechnology • stress resistance (baker’s, brewer’s, wine) • fermentation capacity (baker’s, brewer’s) • flavour ester synthesis (brewer’s) • bioethanol production Fundamental research Nutrient-induced signal transduction in the yeast Saccharomyces cerevisiae and the pathogens Candida glabrata and Candida albicans Biotechnological applications
Nutrient-induced loss of stress resistance The frozen dough process flour water yeast salt mixing dividing moulding freezing storage at -20°C (1 day to 6 months) thawing, proofing and baking
Fermentation-induced loss of stress resistance Glucose cAMP PKA etc. Trehalase activation Induction of ribosomal genes Trehalose mobilization Glycogen mobilization Repression of STRE-controlled genes Rapid drop in general stress resistance
General observation in nature Metabolic activity Stress resistance General question in biotechnology Is high metabolic activity compatible with high stress resistance ? To what extent can stress resistance of living cells be enhanced without compromising metabolic activity ? Initiation of fermentation Fermentation Glucose Sucrose Stress resistance Industrial applications - frozen doughs Baker’s yeast Fermenting yeast: low stress resistance Brewer’s yeast - high-gravity brewing
GOAL Development of yeast strains that maintain a high(er) stress resistance during active fermentation Stress response mechanisms: extensive information Stress resistance determinants: little information Improvement of stress resistance: very little information
I. Prevention of trehalose mobilisation In general : Stress resistance Trehalose content Dry baker’s yeast : Trehalose = ± 15 - 20% of dry weight (± 1-1.5 M in cytosol) Guaranteed to have no significant activity loss during storage for 2 years at room temperature Initiation of fermentation : Glucose Glucose Trehalose Stress resistance Time Time
Trehalose-6-P synthase Trehalose-6-P phosphatase Trehalase (neutral) UDP-Glucose + Glu6P Tre6P Trehalose 2 Glucose TPS2 NTH1 TPS1 Glucose Glucose % Survival (heat shock of 10 min at 52°C) Trehalose (% of dry w) Time (min ) Time (min ) Glucose causes disappearance of other factors required for stress resistance High trehalose levels cannot prevent loss of stress resistance in their absence (Van Dijck et al. 1995 AEM 61, 109-115)
lab strain (heat stress) II. Isolation of ‘fil’ mutants ‘fil’ mutants: deficient in fermentation-induced loss of stress resistance Procedure: EMS-mutagenesis / growth to stationary phase / fermentation for 90 min / (sub)lethal stress treatment (e.g. 30 min at 52°C) / repeat 1 more time and isolation of surviving mutants Glucose Heat shock Stress resistance fil mutant wild type strain Time
1 stat. phase 500 µl stat.. phase 500 µl 2 3 Yeast cells + EMS 50 ml YPD 90 min 30 °C 30 min 52 °C 50 ml YPD 100µl 4 YPD plates 5 50 ml YPD 90 min 30 °C 30’ 52 °C 30’ 54 °C 30’ 56 °C Isolation of fil mutants
Glucose Glucose Glucose 200 100 10 fil1 150 10 8 Ethanol (mmol/mg dry w) % Survival (heat shock of 30 min at 52°C) 6 Growth (OD 600 nm) 100 1 4 50 0.1 wild type 2 0 0 0.01 0 5 10 15 20 0 5 10 15 20 0 30 60 90 Time (h) Time (h) Time (min) Stress resistance Growth Fermentation High(er) stress resistance and high metabolic activity are not incompatible (Van Dijck et al. 2000 JMMB 2, 521-530) fil1 mutant partially inactivating point mutation in adenylate cyclase: Cyr1E1682K
Microarray analysis A number of differentially regulated genes of which 6 are involved in the higher stress tolerance of the fil1 mutant Why is the fil1 mutant more stress tolerant Effect on expression of known targets of the general stress response pathway?
The fil1 mutation is mapped to the catalytic domain of the adenylate cyclase gene resulting in partial inactivation of AC Glucose Adenylate Cyclase Cap Cyr1 cAMP PKA Tps1 Hsp104 Msn2-4 Fermentation Trehalose mobilisation Repression of STRE regulated genes Rapid drop of general STRESS RESISTANCE Growth
The presence of the fil1 mutation enhances heat stress resistance (20’ 51 °C) in strains that lack trehalose or Hsp104 fil1 hxk2 tps1 Survival (%) hxk2 tps1 fil1 hxk2 hsp104 hxk2 hsp104 Time after addition of glucose (min)
Deletion of transcription factors Msn2 and Msn4 in the fil1 mutant does not result in complete loss of heat stress resistance fil1 % survival after 15’ at 51 °C fil1 msn2 msn4 wild type msn2 msn4 Time after the addition of glucose (min)
Because of the existence of compensation effects, (cfr HSP104 expression) it is necessary to compare the presence or absence of the fil1 mutation on the heat stress resistance in a strain that completely lacks trehalose, Hsp104 and the Msn2 and Msn4 transcription factors Construction of MDJ2: W303-1A tps1 hxk2 msn2 msn4 hsp104 fil1 MDJ3: W303-1A tps1 hxk2 msn2 msn4 hsp104
YPD 5 mM H2O2 1.4 M NaCl WT MDJ3 MDJ2 The fil1 mutation strongly increases the heat stress resistance of a strain that lacks trehalose, Hsp104 and all of the stress-regulated Msn2/4 regulated genes 1. On plates after heat-shock at 56 °C MDJ2 (fil1) MDJ3 0’ MDJ2 MDJ3 10’ 30’ MDJ2 MDJ3 60’ MDJ2 MDJ3
The fil1 mutation strongly increases the heat stress resistance of a strain that lacks trehalose, Hsp104 and all of the stress-regulated Msn2/4 regulated genes 2. In liquid medium during the start of fermentation Survival after 30’ at 48 °C (%) Time after addition of glucose (min) MDJ2: tps1 hxk2 msn2 msn4 hsp104 fil1 ade2 MDJ3: tps1 hxk2 msn2 msn4 hsp104 ade2 PVD32: prototrophic W303-1A () () ()
OTHER, UNKOWN fil1 TARGETS?? Hsp26 is very important for the high heat stress tolerance of the fil1 mutant Control cultures W303-1A hsp12 hsp26 fil fil hsp12 fil hsp26 fil1 45 min 56 oC % survival (15 min at 52 °C) WT W303-1A hsp12 hsp26 fil fil hsp12 fil hsp26 fil1 hsp26 fil1 hsp12 hsp12 hsp26 60 min 56 oC Time after addition of glucose (min) W303-1A hsp12 hsp26 fil fil hsp12 fil hsp26 Vianna, submitted
Micro-array analysis between fil1 and wild type (diauxic shift) Wild type Fil1
47 genes were selected after micro-array analysis • 1. Stationary phase cells • 8 differentially expressed genes • 3 confirmed, 2 not confirmed by NB, 3 undetectable • 2. 30 min after addition of glucose to stationary phase cells • 8 differentially expressed genes • 6 confirmed by NB, 2 undetectable • 3. During diauxic shift (glucose to ethanol shift) • 31 differentially expressed genes (24 novel ORF’s) • 20 confirmed, 3 not confirmed by NB, 8 undetectable 27 genes were confirmed by Northern blot analysis
40 30 20 10 0 overexpressed in fil1 YJL160C LEU2 YPL276W SNU13 3 YJL211C YNL179c YGL218w YJR114w YOR041c YNL190W YGL069c SSC3 VMA4 YJR126C YDL023c YCR061W 2 IDH2 1 0 ACH1 TRP1 overexpressed in W303-1A TIS11/CTH2 YER024W HEM13 TRP1 HXT6
fil1 56°C 0’ 30’ 60’ 120’ fil1 srf1∆ fil1 fil1 srf2∆ fil1 / srf1∆ fil1 srf3∆ % survival after heat shock fil1 / srf2∆ fil1 srf4∆ fil1 srf5∆ fil1 / srf3∆ fil1 srf6∆ fil1 / srf4∆ fil1 / srf5∆ fil1 / srf6∆ Time after addition of glucose (min) Deletion of each SRF gene in the fil1 background results in loss of heat stress resistance PROBLEM: ALL THESE GENES OVERLAP WITH OTHER GENES
Introduction of the fil1 point mutation in industrial baker’s yeast strains Despite a lot of effort, NO success
industrial strain III. Isolation of ‘fil’ mutants Strain: commercial tetraploid/aneuploid strain S47 (Lesaffre, Lille) Purpose: freeze-resistant strain for use in frozen dough application Procedure: UV-mutagenesis / growth to stationary phase / preparation of small doughs (0.5g) / fermentation at 30°C for 30 min / freeze/thaw treatment up to 200 times (-30°C/20°C) / solubilization of dough / plating for survivors Results: - many stress-resistant mutants, but most with reduced growth and/or fermentation rate - most promising mutant strain: AT25
120 100 80 Survival (%) 60 40 20 0 0 30 60 90 120 Time (min) Better heat and freeze resistance Freeze resistance (1h -30 °C) Heat resistance (15’ 49°C) Glucose Glucose 120 100 AT25 mutant AT25 mutant 80 Survival (%) 60 40 S47 parent S47 parent 20 0 0 30 60 90 120 Time (min) AT25 mutant seemed to be improved in general stress resistance Teunissen et al., AEM 2002
Much lower proofing-time compared to S47 after deep-freezing of dough to a core temperature of -30 °C 1 4 0 1 3 0 S47 1 2 0 1 1 0 Proof-Time (min) 1 0 0 9 0 AT25 8 0 7 0 0 2 0 4 0 6 0 8 0 1 0 0 T i m e o f s t o r a g e a t - 2 0 ° C ( d a y s ) Teunissen et al., AEM 2002
3 genes consistently upregulated ≥ 3 times + 3 genes consistently downregulated ≤ 3 times in all resistant strains compared to all sensitive strains Confirmed by Northern Individual overexpression (in AT25) or individual deletion (lab strain): little effect IV. Genome-wide expression analysis of ‘fil’ mutants AT25 mutant S47 parent AT25 mutant stress-sensitive strains stress-resistant strains AT25 mutant S47 parent AT25 resistant derivatives S47 sensitive derivatives
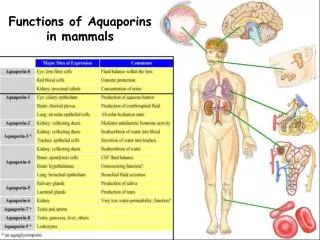
Deletion and overexpression of AQY1 and AQY2 (and human hAQP1): effect on freeze tolerance ? AQY2 HOWEVER: also overexpressed in some resistant strains AQY1 and AQY2 - two water channel encoding genes in yeast - inactive in many lab strains - deletion and overexpression: no clear phenotype - microbial aquaporins: function ?
Freeze tolerance assay. stationary phase cells no glucose glucose = fermenting cells = non-fermenting cells 0°C -30°C glucoseconsumption (plate count) IGC FGC (CFU1) (CFU2) CFU (%) RGC (%) RGC (%) = (FGC/IGC)*100 survival (%) = (CFU1/CFU2)*100 IGC FGC IGC FGC
Overexpression AQY1 or AQY2 in AT25 improves freeze resistance Relative glucose consumption after freezing (RGC) 36% 71% 54% 20% 97% 83% 18 1 day 4°C (IGC) 16 14 1 day -30°C (FGC) 12 mM glucose consumed in 2.5h 10 8 6 4 2 0 AT25 AT25 + AQY2 AT25 + AQY1 AT25 AT25 + AQY2 AT25 + AQY1 laboratory scale yeast industrial pilot scale yeast Same effect with overexpression of human aquaporin gene hAQP1
AT25 (lab scale) AT25 (pilot scale) AT25 + AQY2 (lab scale) AT25 + AQY2 (pilot scale) Overexpression of aquaporins improves maintenance of viability and fermentative activity during freeze storage Overexpression of aquaporins improves freeze tolerance of C. albicans and S. pombe Overexpression of aquaporin in AT25 improves maintenance of viability during freeze storage of small rapidly-frozen doughs 1000 AT25 + AQY2 100 % survival 10 AT25 1 0 10 20 30 40 50 60 70 80 90 100 number of days
Case study (6). Overexpression of AQY1-1 and AQY2-1 enhances freezetolerance of industrial strains. non-frozen control non-frozen control frozen frozen Scientists@work 2006. wild type baker’s yeast (AT25) GM baker’s yeast (AT25 + AQY2-1) Other commercially important characteristics not affected. Tanghe et al., 2002.
large doughs, core T° -30°C Proofing time. Gassing power. 120 1000 110 900 100 800 90 proofing time (min at 35°C) gassing power (ml in 2 h) 700 80 70 600 60 500 50 400 0 20 40 60 80 100 120 140 0 20 40 60 80 100 120 140 frozen storage duration (days) frozen storage duration (days) AT25/TPI1p AT25/TPI1p AQY1-1 AT25/TPI1p AQY2-1 Case study (7). Aquaporin overexpression does notimprove yeast freezetolerance when cultured and tested in industrial conditions. laboratory versus industrial conditions: many ≠ parameters - culturing conditions? NO - thawing conditions? NO - freezing conditions? YES Tanghe et al., 2004.
survival in cell suspensions 120 100 80 60 %RGC 40 20 0 N2,l DPL EtOH -30°C DPL freezer -30°C DPL N2,l SPL EtOH -30°C SPL freezer -30°C SPL AT25+pAQY2-1 BY4743+pAQY2-1 AT25+vector BY4743+vector Aquaporin-mediated improvement of freeze tolerance is restricted to fast freezing conditions. laboratory strain BG industrial strain BG
25 20 15 10 large, industrial dough 5 temperature (°C) 0 2 4 6 8 10 -5 -10 -15 -20 -25 time (minutes) EtOH -30°C SPL EtOH -30°C DPL freezer -30°C DPL freezer -30°C SPL Temperature evolution during freezing.
Aquaporin-mediated improvement of freeze tolerance is restricted to fast freezing conditions. survival in small doughs Fast freezing (EtOH -30°C). Slow freezing (freezer -30°C). 1000 1000 100 100 survial (% CFU) survial (% CFU) 10 10 1 0 10 20 30 40 50 60 70 80 90 100 0 10 20 30 40 50 60 70 80 90 100 frozen storage duration (days) frozen storage duration (days) LAT25 LAT25 AT25 AT25 AT25/AQY2-1 LAT25/AQY2-1 AT25/AQY2-1 LAT25/AQY2-1
slow freezing fast freezing IC icecrystal formation water outflow damage to cellorganelsand plasma membrane cellular dehydration critical cooling rate dependent on cell type - S/V ratio - water permeability survival ↓ survival ↓ ↓ ↓ Hypothesis. EC freezing IC supercooling water permeability limiting? aquaporin overexpression advantageous ? chemical gradient for free water = unstable situation
Underlying mechanism FREEZING aquaporin high low Extracellular medium freezes first Intracellular medium freezes later Osmolarity Aquaporins play a function in freeze tolerance of yeast First clear function for microbial aquaporins Only with rapid freezing Slow freezing: no effect (Larger commercial doughs: no effect unfortunately)
Osmotic gradient Osmotic gradient Less intracellular ice crystal formation Rapid freezing H2O H2O Slow freezing Lower drop in viability Higher expression of aquaporins allows faster efflux of water FREEZING H2O H2O Extracellular medium frozen - Intracellular medium not frozen
Acknowledgments Dr. Sonia Colombo Dr. Barbara Leyman Dr. Matthias Versele Dr. An Tanghe Cristina Roscoe Vianna Prof. Johan Thevelein
Stress resistance in general is a multifactorial trait. Development of a novel technology to determine the all genes involved in a certain phenotype in one go. = AMTEM
Yeast strain with 600 specific, artificial oligonucleotide markers spread evenly throughout the genome
First technology allowing simultaneous identification of genes involved in polygenic traits Many novel advanced applications