Download
1 / 9
120 likes | 411 Views
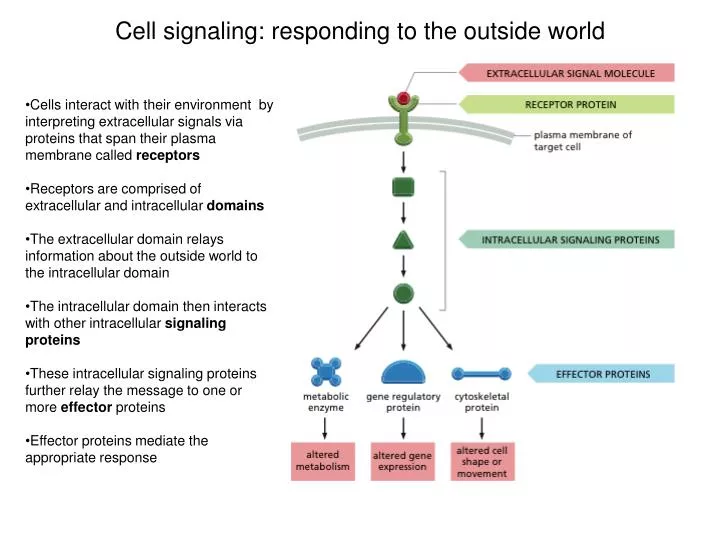
Cell signaling: responding to the outside world. Cells interact with their environment by interpreting extracellular signals via proteins that span their plasma membrane called receptors Receptors are comprised of extracellular and intracellular domains

E N D
Cell signaling: responding to the outside world • Cells interact with their environment by interpreting extracellular signals via proteins that span their plasma membrane called receptors • Receptors are comprised of extracellular and intracellular domains • The extracellular domain relays information about the outside world to the intracellular domain • The intracellular domain then interacts with other intracellular signalingproteins • These intracellular signaling proteins further relay the message to one or more effector proteins • Effector proteins mediate the appropriate response
Receiving the Signal: G-protein Coupled Receptors (GPCRs) • GPCRs are an important and ubiquitous class of eukaryotic receptors (>700 in humans) • The extracellular domain connects to the intracellular domain through 7 transmembrane spans • The intracellular domain is coupled to a heterotrimeric G-protein • The heterotrimeric g-protein is composed of 3 subunits: G, G, and G • When the G subunit is bound to GDP it is “OFF”; when it is bound to GTP it is “ON” • When the extracellular domain binds to the signal molecule, it causes a conformational change relayed through the transmembrane spans to the intracellular domain • The conformational change relayed to the intracellular domain causes the G subunit to release GDP and bind to GTP thereby activating both the G and G/G subunits
P P P Kinase 1 Kinase 2 Kinase 3 Phosphorylation Cascade Transmitting the Signal: Protein Kinases • Activated receptors frequently transmit signals through through intracellular signaling proteins called kinases • Protein kinases are enzymes that add a phosphate group from ATP onto a substrate protein; this reaction is called phosphorylation • Phosphorylation frequently serves to activate the substrate of the kinase, but can also target the substrate for degradation • Kinases are often themselves activated by other kinases via phosphorylation and can organize into phosphorylation cascades • One important class of phosphorylation cascade is called a mitogen activated protein kinase (MAPK) cascade
Effector protein Responding to the Signal: Effector Proteins • The final step in cell signaling is activation of the effector proteins • The effector proteins carry out the cellular response to the signal • Often the cellular response involves expression of previously inactive genes which requires effector proteins called transcriptional activators or transcription factors • Transcription factors are proteins that bind to specific DNA sequences called promoters that are upstream of the genes that are turned on • Promoters that are upstream of genes that are only activated during specific cellular responses are called response elements • Effector proteins can also directly act on proteins that regulate cell shape to induce changes in morphology by rearranging the cytoskeleton • Other types of effector proteins directly regulate cell growth by arresting the cell cycle or altering cellular metabolism Changes in gene expression Effector Protein Cytoskeletal Rearrangement Cell Cycle Arrest
A model signaling pathway: The Yeast Pheromone Pathway • There are two mating types (“sexes”) of yeast, a and (in the lab we generally study the a mating type) • They can mate by responding to an extracellular signal, called a pheromone (13 amino acid peptide), released by one mating type and received by the other • The mating type pheromone, alpha factor, binds to a GPCR on the surface of an a cell to initiate signaling • The GPCR undergoes a conformational change that is transmitted to the G-protein whose G subunit releases GDP and binds to GTP • The GTP-bound G subunit then dissociates from the G/G subunit which in turn initiates a MAPK phosphorylation cascade where a MAP kinase kinase kinase (MAPKKK) activates a MAP kinase kinase (MAPKK) which activates a MAP kinase (MAPK) • The activated MAPK then activates several effector proteins: a transcription factor and a cell-cycle inhibitor • The net results are cell cycle arrest, cytoskeletal rearrangements to “grow” toward where the pheromone originated (in hopes of mating successfully), and expression of genes required for fusion to the opposite mating type a cells, no pheromone + mating pheromone (alpha factor) a cells, + pheromone
Other MAPK Signaling Pathways in Yeast Filamentation Pathway HOG Pathway Pheromone Pathway • In addition to the pheromone pathway, yeast have several other pathways that use the MAPK architecture to transmit signals • Two other commonly studies MAPK pathways in yeast are the High Osmolarity Glycerol pathway (HOG pathway), which responds when there is high salt in the environment, and the filamentation pathway which responds to lack of nitrogen in the environment • These pathways and the pheromone pathway share some components • How do these pathways keep their prevent cross-talk and maintain signal specificity? Nitrogen Starvation Pheromone
Ste11 Ste11 Ste5 Pbs2 Ste7 Fus3 Hog1 Achieving Specificity in Signaling in Yeast MAPK Pathways • With so many components in common, how do yeast cells keep their signals straight? • Two mechanisms yeast employ to achieve signaling specificity are scaffolding and cross-pathway inhibition • Scaffolds: the pheromone pathway uses the scaffold Ste5, and the HOG pathway uses the scaffold/MAPKK Pbs2 • Scaffolds promote signaling efficiency by localizing all the proteins in one cascade close together • Scaffolds promote signaling specificity by preventing upstream activators (e.g. MAPKKK or MAPK) from interacting with inappropriate downstream proteins (e.g. the wrong MAPK or effector) • Cross-pathway inhibition promotes signaling specificity by having the activated pathway make sure the other pathway stays off by actively inhibiting it Scaffolds in the pheromone and HOG pathways Cross-pathway inhibition of the filamentation pathway by the pheromone pathway: Fus3 phosphorylates and triggers degrad-ation of Tec1, the transcription factor required for filimentation
Engineering Cross-Talk: Rewiring Yeast MAPK Pathways • If scaffolds promote specificity, it should be possible to rewire pathways by engineering scaffolds with new connections • Goal: link the pheromone pathway to the HOG pathway, so when you add pheromone, you induce the salt survival response • Step 1: Fuse the pheromone scaffold, Ste5, to the HOG pathway scaffold, Pbs2 • Step 2: Remove the binding site in the Ste5 for the pheromone MAPKK (Ste7) • Step 3: Remove the connection between the Pbs2 and the upstream salt response proteins • Now pheromone induces the HOG response The theory: The construct: The result:
References • Alberts et al. Molecular Biology of the Cell, Chapter 15 • Dohlman, H. and Thorner, J. Regulation of G-Protein initiated signal transduction in yeast: paradigms and principles. Annu. Rev. Biochem. 2001. 70:703–54 • Bao et al. Pheromone-dependent destruction of the Tec1 transcription factor is required for MAP kinase signaling specficity in yeast. Cell. 2004. 119: 991 • Schwartz and Madhani. Principles of MAP kinase signaling specificity in Saccharomyces cerevisiae. Annu. Rev. Genet. 2004. 38: 725 • Park, Zarrinpar and Lim. Rewiring MAP kinase pathways using alternative scaffold assembly mechanisms. Science 2003. 299:1061