Download
1 / 1
10 likes | 115 Views
(B). (A). Characterization of Different Modes of Ca 2+ Uptake under Different Physiological Conditions in the Heart Mitochondria. Shivendra G. Tewari 1 , Ranjan K. Pradhan 1,2 , Jason N. Bazil 1,2 , Amadou K.S. Camara 3 , David F. Stowe 2,3 , Daniel A. Beard 1,2 , and Ranjan K. Dash 1,2

E N D
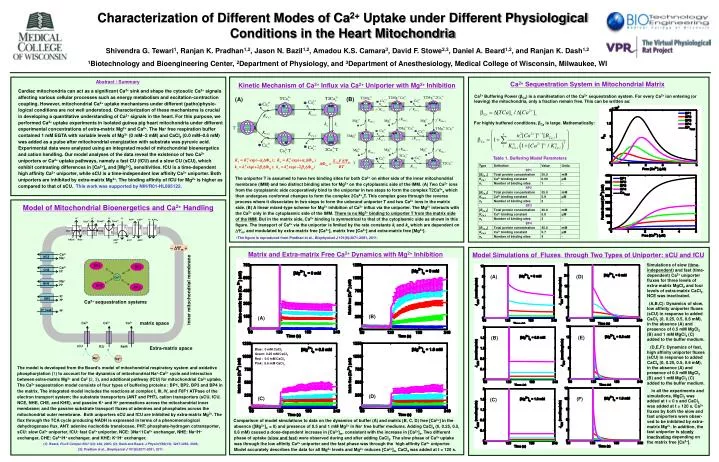
(B) (A) Characterization of Different Modes of Ca2+ Uptake under Different Physiological Conditions in the Heart Mitochondria Shivendra G. Tewari1, Ranjan K. Pradhan1,2, Jason N. Bazil1,2, Amadou K.S. Camara3, David F. Stowe2,3, Daniel A. Beard1,2, and Ranjan K. Dash1,2 1Biotechnology and Bioengineering Center, 2Department of Physiology, and 3Department of Anesthesiology, Medical College of Wisconsin, Milwaukee, WI Abstract / Summary Cardiac mitochondria can act as a significant Ca2+ sink and shape the cytosolic Ca2+ signals affecting various cellular processes such as energy metabolism and excitation-contraction coupling. However, mitochondrial Ca2+ uptake mechanisms under different (patho)physio-logical conditions are not well understood. Characterization of these mechanisms is crucial in developing a quantitative understanding of Ca2+ signals in the heart. For this purpose, we performed Ca2+ uptake experiments in isolated guinea pig heart mitochondria under different experimental concentrations of extra-matrix Mg2+ and Ca2+. The Na+ free respiration buffer contained 1 mM EGTA with variable levels of Mg2+ (0 mM–2 mM) and CaCl2 (0.0 mM–0.6 mM) was added as a pulse after mitochondrial energization with substrate was pyruvic acid. Experimental data were analyzed using an integrated model of mitochondrial bioenergetics and cation handling. Our model analyses of the data reveal the existence of two Ca2+ uniporters or Ca2+ uptake pathways, namely a fast CU (fCU) and a slow CU (sCU), which exhibit contrasting differences in [Ca2+]e and [Mg2+]e sensitivities. fCU is a time-dependent high affinity Ca2+ uniporter, while sCU is a time-independent low affinity Ca2+ uniporter. Both uniporters are inhibited by extra-matrix Mg2+. The binding affinity of fCU for Mg2+ is higher as compared to that of sCU. This work was supported by NIH/R01-HL095122. Ca2+ Sequestration System in Mitochondrial Matrix Kinetic Mechanism of Ca2+ Influx via Ca2+ Uniporter with Mg2+ Inhibition Ca2+ Buffering Power (βCa) is a manifestation of the Ca2+ sequestration system. For every Ca2+ ion entering (or leaving) the mitochondria, only a fraction remain free. This can be written as: For highly buffered conditions, βCa is large. Mathematically: Table 1. Buffering Model Parameters The uniporter T is assumed to have two binding sites for both Ca2+ on either side of the inner mitochondrial membrane (IMM) and two distinct binding sites for Mg2+ on the cytoplasmic side of the IMM. (A) Two Ca2+ ions from the cytoplasmic side cooperatively bind to the uniporter in two steps to form the complex T2Ca2+e which then undergoes conformal changes to form the complex 2Ca2+xT. This complex goes through the reverse process where it dissociates in two steps to form the unbound uniporter T and two Ca2+ ions in the matrix side. (B) A linear mixed-type scheme for Mg2+ inhibition of Ca2+ influx via the uniporter. The Mg2+ interacts with the Ca2+ only in the cytoplasmic side of the IMM. There is no Mg2+ binding to uniporter T from the matrix side of the IMM.But in the matrix side, Ca2+ binding is symmetrical to that of the cytoplasmic side as shown in this figure. The transport of Ca2+ via the uniporter is limited by the rate constants ki and ko which are dependent on m, and modulated by extra-matrix free [Ca2+], matrix free [Ca2+] and extra-matrix free [Mg2+]. The figure is reproduced from Pradhan et al., Biophysical J 101(9):2071-2081, 2011. Model of Mitochondrial Bioenergetics and Ca2+ Handling Matrix and Extra-matrix Free Ca2+ Dynamics with Mg2+ Inhibition Model Simulations of Fluxes through Two Types of Uniporter: sCU and fCU Simulations of slow (time-independent) and fast (time-dependent) Ca2+ uniporter fluxes for three levels of extra-matrix MgCl2 and four levels of extra-matrix CaCl2. NCE was inactivated. (A,B,C): Dynamics of slow, low affinity uniporter fluxes (sCU) in response to added CaCl2 (0, 0.25, 0.5, 0.6 mM), in the absence (A) and presence of 0.5 mM MgCl2 (B) and 1 mM MgCl2 (C) added to the buffer medium. (D,E,F): Dynamics of fast, high affinity uniporter fluxes (sCU) in response to added CaCl2 (0, 0.25, 0.5, 0.6 mM), in the absence (A) and presence of 0.5 mM MgCl2 (B) and 1 mM MgCl2 (C) added to the buffer medium. In all the experiments and simulations, MgCl2 was added at t = 0 s and CaCl2 was added at t = 120 s. Ca2+ fluxes by both the slow and fast uniporters were obser-ved to be inhibited by extra-matrix Mg2+. In addition, the fast uniporter is slowly inactivating depending on the matrix free [Ca2+]. (A) (D) (B) (A) (E) (B) Blue: 0 mM CaCl2 Green: 0.25 mM CaCl2 Red : 0.5 mM CaCl2 Pink: 0.6 mM CaCl2 The model is developed from the Beard’s model of mitochondrial respiratory system and oxidative phosphorylation (1) to account for the dynamics of mitochondrial Na+-Ca2+ cycle and interaction between extra-matrix Mg2+ and Ca2 (2, 3), and additional pathway (fCU) for mitochondrial Ca2+ uptake. The Ca2+ sequestration model consists of four types of buffering proteins : BP1, BP2, BP3 and BP4 in the matrix. The integrated model includes the reactions at complex I, III, IV, and F0F1 ATPase of the electron transport system; the substrate transporters (ANT and PHT), cation transporters (sCU, fCU, NCE, NHE, CHE, and KHE), and passive K+ and H+ permeations across the mitochondrial inner membrane; and the passive substrate transport fluxes of adenines and phosphates across the mitochondrial outermembrane. Both uniporters sCU and fCU are inhibited by extra-matrix Mg2+.The flux through the TCA cycle producing NADH is expressed in terms of a phenomenological dehydrogenase flux. ANT: adenine nucleotide translocase, PHT: phosphate-hydrogen cotransporter, sCU: slow Ca2+ uniporter, fCU: fast Ca2+ uniporter, NCE: 3Na+/1Ca2+ exchanger, NHE: Na+/H+ exchanger, CHE: Ca2+/H+ exchanger, and KHE: K+/H+ exchanger. (1): Beard, PLoS Comput Biol 1(4): e36, 2005; (2): Dash and Beard, J Physiol 586(13): 3267-3285, 2008; (3): Pradhan et al., Biophysical J 101(9):2071-2081, 2011. (D) (F) (C) (C) Comparison of model simulations to data on the dynamics of buffer (A) and matrix (B, C, D) free [Ca2+] in the absence ([Mg2+]e= 0) and presence of 0.5 and 1 mM Mg2+ in Na+ free buffer mediums. Adding CaCl2 (0, 0.25, 0.5, 0.6 mM) caused a dose-dependent increase in [Ca2+]m, consistent with the increase in [Ca2+]e. Two different phase of uptake (slow and fast) were observed during and after adding CaCl2. The slow phase of Ca2+ uptake was through the low affinity Ca2+ uniporter and the fast phase was through the high affinity Ca2+ uniporter. Model accurately describes the data for all Mg2+ levels and Mg2+ reduces [Ca2+]m. CaCl2 was added at t = 120 s.
















