Download
1 / 40
400 likes | 464 Views
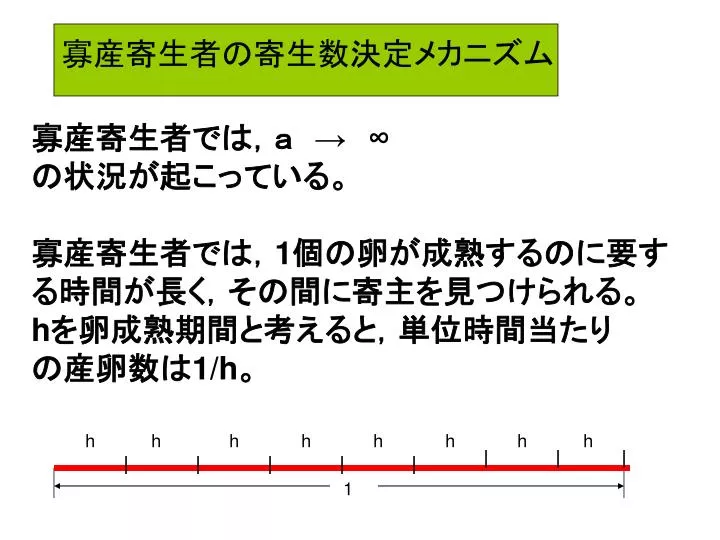
寡産寄生者の寄生数決定メカニズム. 寡産寄生者では,a → ∞ の状況が起こっている。 寡産寄生者では, 1 個の卵が成熟するのに要す る時間が長く,その間に寄主を見つけられる。 h を卵成熟期間と考えると,単位時間当たり の産卵数は 1/h 。. h. h. h. h. h. h. h. h. 1. 寡産寄生者の寄生数決定メカニズム. f (y) = ay / (1 + ahy ) (4.9) (Holling’s disc equation). (4.9) で a → ∞ とおき

E N D
寡産寄生者の寄生数決定メカニズム 寡産寄生者では,a → ∞ の状況が起こっている。 寡産寄生者では,1個の卵が成熟するのに要す る時間が長く,その間に寄主を見つけられる。 hを卵成熟期間と考えると,単位時間当たり の産卵数は1/h。 h h h h h h h h 1
寡産寄生者の寄生数決定メカニズム f(y) = ay / (1 + ahy) (4.9) (Holling’s disc equation) (4.9)でa → ∞とおき f(y) = 1/h (4.12) よって Z = y{1 - exp(- x t / h / y)} (4.13) = y{1 - exp(- x f / y)} Thompson's (1924) model (f = t / h = 産卵数) (4.14)
イラガの繭 イラガイツツバセイボウ
Griffiths' (1977) model -δln(1 - p) + (1 - δ)p = x f / y(4.15) (δ:捕食寄生者が既寄生寄主に出会ったとき産卵する確率) xf/y: 寄主当たり産卵数、f;1メスあたり産卵数 R = x/y (4.15)と(4.16)から,寄生率は (4.17)a =10A=100.246,b= B+1=0.392 -δ ln(1 - p) + (1 - δ)p = aRb
系の安定をもたらすモデル a.寄主側の世代増殖率の改変 寄主の増殖率r 一定 → 密度依存 b.寄生者間の干渉 寄主発見効率(寄主発見面積)a一定 → 寄生者の密度増大に 伴って低下 Hassell & Varley (1969) のモデル a=Qx-m 対数
c.非ランダムな探索 寄主が寄生される確率は一定 → 一部の寄主は,寄生を受けにくいこと イラガの繭の例 地面近くの繭は寄生されにくい 遅く出現した繭は寄生されにくい May( 1978)のモデル ポアソン分布のゼロ項 exp(-axi)→ 負の二項分布のゼロ項 (1+axi /k)- k
5.野外における捕食者(寄生者)と被食者(寄主)との個体群動態5.野外における捕食者(寄生者)と被食者(寄主)との個体群動態 昆虫の生命表63のうち,捕食,寄生,病気による死亡が密度依存性を示した例は13(Stiling 1988)。そのうち,変動主要因となった例は,3例である。ただし,そのうちの2例は解析上問題があるし,残りの1例も調査期間はわずか3年にしか過ぎない 。 これまでの野外調査で、捕食者(寄生者)が被食者個体群を制御していることが示されたことはほとんどない。
演習4(2点).Nicholson-Bailey model ((4.3)と(4.4))を使って,a = 0.001, r = 2, x1 = 300, y1 = 1000 のとき,第1世代から第50世代までのx, yの変化をグラフで表わせ。 演習5(2点).Nicholson-Bailey model ((4.3)と(4.4))のrを= 10*exp(-0.001*yn-1exp(-xn-1*0.001)(寄生後の寄主密度に対して密度依存要因が働く)に置き換え,a = 0.001, x1 = 300, y1 = 1000 のとき,第1世代から第50世代までのx, yの変化をグラフで表わせ。
演習5(2点).Hassell & Varley (1969)のモデルで,Q = 0.001, r = 2, x1 = 300, y1 = 1000, m=0.5のとき,第1世代から第50世代までのx, yの変化をグラフで表わせ。 演習6(2点).May(1978)のモデルでa = 0.001, r = 2, x1 = 300, y1 = 1000, k=0.5のとき,第1世代から第50世代までのx, yの変化をグラフで表わせ。 exp(-axi)→exp(-Qxi1-m) ポアソン分布のゼロ項 exp(-axi)→ 負の二項分布のゼロ項 (1+axi /k)- k
パッチ間の移動に時間がかからない場合のパッチ利用戦略パッチ間の移動に時間がかからない場合のパッチ利用戦略 捕食: Z = y(1 - exp{-a(xt - hZ)}(4.10) をtについて解く。捕食者は1頭とする。 Ni = パッチiにおける初期被食者密度 Mi = パッチiにおける捕食を免れた被食者密度
T一定の条件下で、Oが最大値を取る時のMiの値をラグランジェの乗数法により求める j = 1,2,3,…,n (5.5) 各パッチの密度が同じになるまでパッチに留まって 捕食をすればよい
多産寄生者のパッチ利用戦略 時間当たりの未寄生寄主との遭遇頻度を各パッチで同じにするように各パッチを訪問すべきである。 寡産寄生者のパッチ利用戦略 各パッチの寄生率を一定に保つようにパッチを移動すべきある(Yamada 1988)。過寄生によって失われる卵数を最小にする。
寄生蜂(Venturia canescens)のパッチ利用戦略 理論値 パッチ滞在時間 寄生数 観察値
寄主遭遇頻度 最初 最終離脱時 多産寄生者でも、状況によっては卵枯渇に遭遇する。->寡産寄生者の戦略を採る
演習7(18点) まず2人でペアを組みなさい。ほぼ同じ大きさの紙切れ(あるいは,碁石,ビーズ,大豆等)を 450枚と中が見えない袋を2つ用意せよ。200枚の紙切れに鉛筆等でx印を入れよ。2人のうちどちらが試験者と被験者になるかを決めよ。試験者は,一方の袋に印つき紙切れをa枚,印なし紙切れを200-a枚,残りの袋に,印つき,b枚,印なし,200-b枚を入れよ。a,bとしては,60,80,100,120,140から選べ。a=bとしてもよい。試験者は被験者に,2つの袋の中に印つき紙と印なし紙が入っていて,印つき枚数は,60,80,100,120,140のどれかで,そのどれになるかはランダムであることを述べる。しかし,試験者はa,bとして何を選んだか,どちらの袋に多くの印つき紙切れが入っているかを被験者に教えてはいけない。被験者は,なるべく多くの印つき紙切れを取れるよう2つの袋から50回紙切れを取りなさい。取り出した紙は,印つきの場合は印無しの紙切れに交換し袋の中に戻し、印無しの場合はそのまま戻す。1回の試行で1枚の紙切れを取り2枚以上取らない。1回の試行ごとにどちらの袋から取るかは自由に選べる。試験者は,被験者がどちらの袋を選んだかと,取られた紙に印があるかどうかを記録せよ。これをa,bの値を変えて合計3回行え。その結果をまとめるとともに,理論的な最適戦略を明記して,それと比較して,被験者が最適に振る舞っているかどうかを考えよ。最適戦略は全知(袋の中にある印つき紙数を印なし紙数を知っている)を仮定したときと、そうでないとき(ただし,袋のなかにある印つき紙の数は,60、80,100,120,140枚のいずれかであることは知っている)の両方を考えよ。必ず,横軸に試行回数,縦軸に袋に残った印つき紙の枚数をとり,2つの袋それぞれについて,袋に残った印つき枚数の変化を一つのグラフ上に描け。そのグラフ上でいつ袋を変えたかも示せ。また,2つの袋からの試行回数と印つき紙を引いた回数を記せ.入っている印つき枚数をxとするとき,y回試行して,得る印つき紙数はランダムに引いたとすると(1-exp(-y/200))x となる.これも計算し、実際引いた印つき紙数と比較せよ.これらを基に考察せよ。
1個ごとに袋を選択し1個ずつ計50個を取る。 色つきがでたら,色なしで置き換える。 計200個 色つき a 計200個 色つき b a, b = 60, 80, 100, 120, 140 被験者には,a,bの値を言ってはいけないが, 60,80,100,120,140のうちの一つであることは知らせる
入っている印つき枚数をxとするとき,y回試行して,得る印つき紙数はランダムに引いたとすると(1-exp(-y/200))x となる.これも計算し、実際引いた印つき紙数と比較せよ.
全知を仮定したときの最適戦略? 全知を仮定しないときの最適戦略?
パッチ間の移動に時間が かかる場合の最適滞在時間 Charnov(1976)の限界値定理 (marginal-value theorem) パッチは無限にあると仮定 同じ質のパッチを仮定 パッチ間移動にかかる時間は一定
移動時間は常に一定であり,パッチの消費によって利用可能なパッチの数が減り,移動時間が長くなるようなことは考えてない。移動時間は常に一定であり,パッチの消費によって利用可能なパッチの数が減り,移動時間が長くなるようなことは考えてない。 フンバエの交尾時間(Parker 1978),フタモンアシナガバチの水場での滞在時間(吸水時間)(Kasuya 1982)がこの限界値定理によってよく説明される
餌メニュー選択 質の異なる餌のうちどれを選ぶべきか f = ats(N1+ N2) (E1/ h1 > E2/ h2) 1 = ts + h1atsN1 + h2atsN2 = ts(1+ h1aN1 + h2aN2) それゆえ,単位時間あたりのエネルギ-獲得R1+2は ,
同様にして,捕食者が1種類のみを捕食するときの単位時間あたりのエネルギ-獲得は,同様にして,捕食者が1種類のみを捕食するときの単位時間あたりのエネルギ-獲得は,
エネルギ-獲得率を最大にするためには,以下の条件のときに2種の餌を食べるべきである。エネルギ-獲得率を最大にするためには,以下の条件のときに2種の餌を食べるべきである。 R1+2 > R1(1) R1+2 > R2(2) A1= R1+2 - R1,A2=R1+2 - R2とおき整理すると 正の時、2種 負の時、1種だけ E1/ h1 > E2/ h2なので 常に成立。
ある確率pで捕獲する戦略を 考える pで偏微分して,
E2+λ1h1E2 - λ1h2E1 > 0 (変形するとE2/h2 > λ1E1/(1+λ1h1))のとき p = 1でR1+2Pは最大となる。 E2/h2 < λ1E1/(1+λ1h1) のとき, p = 0 でR1+2pは最大となる。 劣位の餌を部分的に取る戦略は最適でない
(λi = aNi) ○価値の低い餌を部分的に採る戦略は存在しない。 ○あるタイプの餌をメニュ-に加えるかどうかは,その餌タイプのE/hの値と上位の餌タイプのE,h,Nだけで決まり,対象餌タイプの密度は関係がない。
推定遭遇頻度: 選択なしに選んだときのメニュー 大きいほうが遠くにいても気付きやすい.低密度では,大きい餌の密度は20で,小さい餌の密度はそれより増やして,視野に入りやすさを一定にした Lは20 ハンドリングタイムは同じ
なぜ劣位の餌を取るのか? ○捕食者は,価値の高い餌と低い餌を見分ける完全な能力を持っていない。つまり,正確なE,hを知ることが出来ない。 ○餌密度を正確に知ることが出来ない。つまり,正確なλを知ることが出来ない。 ○E,hは一定でなく,確率変数であるかもしれない.個体によってE,hが異なっているかも知れないし,同じ個体でも,生理状況によって,一定でないかもしれない. ○餌の価値を認識するのに時間がいるのかもしれない
〇短い時間当たりのエネルギ-獲得率を高めているのでないのかも?〇短い時間当たりのエネルギ-獲得率を高めているのでないのかも? ○栄養素のバランスを第1に考えて餌選択をしているのかもしれない。 ○あるタイプの餌は,捕獲するのに労力がかかりその後の寿命に影響を与えたり,捕獲中に怪我をしたりすることが多いかもしれない。
その他の注意点 寡産寄生者にとって寄主の価値は,E(この場合は,子の生存率と成虫時の大きさ,発育期間)によってのみ決まり、ハンドリングタイムは関係ない.卵成熟期間にかならず価値が高い寄主に出会う保証があれば,価値が高い寄主のみを選ぶのが最適戦略となる。
多産寄生者において、将来、卵枯渇に遭遇する可能性が高いときは、寄主密度が低いときにも、価値の低い餌に対しては、100%受け入れないことが最適となる(Sirot et al., 1997)。 言い換えると,単位時間当たり得る適応度上の利益を最大にしているのではなく,一生涯から得る利益を最大にするよう振る舞っている可能がある。この場合は,寄主の価値に応じた受入率が最適となる(Plantegenest et al., 2004).
Rules of thumb(おおざっぱなルール) 探索生物は全知全能ではない。 単純なル-ル(Rules of thumb)にしたがっているだけかも。 例えば,giving-up time ruleに従っているに過ぎないかもしれない。
パッチ脱出 餌 餌 餌 giving-up time
