Download
1 / 30
330 likes | 601 Views
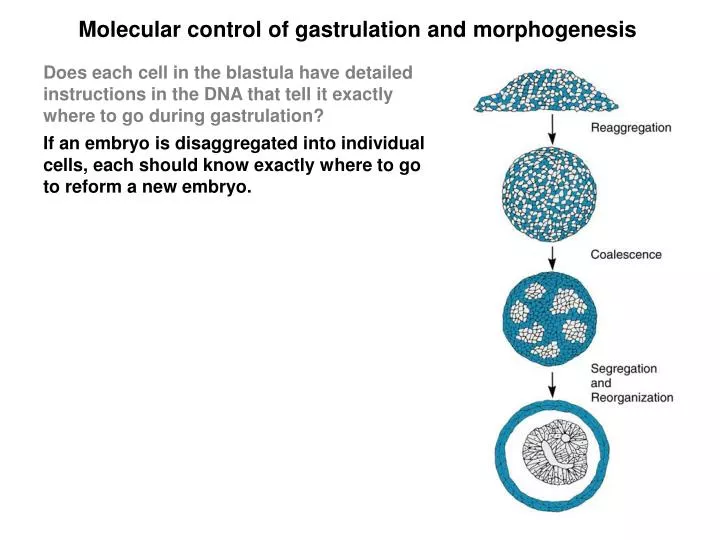
Molecular control of gastrulation and morphogenesis. Does each cell in the blastula have detailed instructions in the DNA that tell it exactly where to go during gastrulation?. If an embryo is disaggregated into individual cells, each should know exactly where to go to reform a new embryo. .

E N D
Molecular control of gastrulation and morphogenesis Does each cell in the blastula have detailed instructions in the DNA that tell it exactly where to go during gastrulation? If an embryo is disaggregated into individual cells, each should know exactly where to go to reform a new embryo.
Molecular control of gastrulation and morphogenesis Does each cell in the blastula have detailed instructions in the DNA that tell it exactly where to go during gastrulation? If an embryo is disaggregated into individual cells, each should know exactly where to go to reform a new embryo. When this experiment is performed, a degree of reorganization occurs, but it is not complete. Embryoids: are slightly similar to embryos but they lack the real organization. Conclusions: 1. Genes impart only partial instructions for assembly of the embryo 2. Like cells all stick together, revealing distinct adhesive properties. 3. The relative positions of aggregates reflect the relative positions in the embryo (skin outside, heart inside).
Cell adhesion is the driving force in gastrulation When cells from an embryo are disaggregated and recombined, they can be readily ranked according to their ability to form the central portion. (chondrocytes > heart cells > liver cells is the hierarchical order) Why?
Cell adhesion is the driving force in gastrulation When cells from an embryo are disaggregated and recombined, they can be readily ranked according to their ability to form the central portion. (chondrocytes > heart cells > liver cells is the hierarchical order) Why? Differential adhesion hypothesis: the cell type with maximal adhesiveness (chondrocytes) will form a core that is surrounded by concentric spheres of cells with progressively lower adhesiveness. Cell adhesion can be measured by the ‘pancake test’. When aggregates of different cell types are subjected to a flattening force (centrifugation to induce a centrifugal force), the cells that adhere most tightly form a ball, while those that adhere more loosely form a flatter, pancake structure. Cell adhesion is a major factor that regulates aggregation of like cells and controls position during morphogenesis. What regulates how tightly or loosely cells attach?
Cells adhere by cell junctions, cell adhesion molecules, or substrate adhesion molecules Cell junctions:large, complex structures that form slowly but generate very strong and durable connections (tight junctions, desmosomes, and gap junctions). Cell adhesion molecules (CAMs):single molecules that traverse cell membranes and allow cells to adhere to one another. Adhesions form quickly, they are selective, but they are relatively weak in comparison to cell junctions. Substrate adhesion molecules (SAMs): a group that consists of extracellular matrix molecules and matched receptors that are expressed on the cell surface.
3 types of specialized attachments hold embryo cells together Tight junctions are regions where membranes of adjacent cells actually fuse. They encircle the whole cell and provide a barrier for leakage between cells (common in the GI tract to prevent leakage of food out of the gut). The also prevent membrane proteins from moving freely from apical to basal regions. Desmosomes are spot rivets that weld cells together. A cytoplasmic plaque is connected by inter cellular protein filaments, and the plaques are connected to the intermediate filaments within the cytoplasm (common in cells that are stretched a lot such as skin). Gap junctions are small channels between cells that allow for intercellular communication. They are found in smooth muscle and allow signals (small ions) to spread between cells so that all muscle cells contract as one.
CAMs firmly anchor adjacent cells to the cytoskeleton Cell adhesion molecules (CAMs) are glycoproteins with 3 major domains: The extracellular domain allows one CAM to bind to another on an adjacent cell. The binding can be to the same type of cell (homotypic) or to a different cell type (heterotypic). The transmembrane domain links the CAM to the plasma membrane through hydrophobic forces. The cytoplasmic domain is directly connected to the cytoskeleton by linker proteins. This anchoring is important to prevent lateral diffusion of adhesion molecules in the membrane. Three major types of CAMs are immuno globulin-like CAMs, cadherins, and lectins.
Neural cell adhesion molecule is typical of immunoglobulin (IgG)-like CAMs N-CAM was one of the first to be discovered. The extracellular domain has IgG like repeats that are thought to allow binding to other N-CAMs by interdigitation between loops. Insects have IgG CAMs but no IgG. Thus, IgGs may have evolved from IgG-like CAMs. Polysialic acid region (PSA): 3 long carbohydrate chains with negative charge are attached to the 5th loop. The overall charge varies on different N-CAMs. Large PSA regions induce a large negative charge which repels cells (embryonic cells during gastrulation). Small PSA regions allow attachment due to low charges, and these are common on adult cells. N-CAMs have isoforms: one N-CAM gene can generate over 100 different molecules by post transcriptional and post translational modification. The most common are 120, 140, and 180 kD.
Cadherins mediate calcium-dependent cell adhesion Cadherins are the most prevalent CAMs in vertebrates. They are rapidly degraded by proteases in the absenceof Ca++. There are 4 major types: E cadherins in epithelial cells P cadherins in placenta N cadherins in neural tissue L cadherins in liver Each associates with its own type. 125 kD transmembrane glycoproteins that bind homotypically using the first 113 AA The differences in cadherin expression are responsible for the differential adhesiveness seen in disaggregated tissue. Cells that express more cadherin = tissues that form a ball in the center of cell aggregates. Catenins are proteins that link cadherins to the cytoskeleton. If this linkage is disturbed, cadherins do not work, and embryonic development is disrupted (disrupt catenins in neural tissue = brain forms improperly).
Lectins bind heterotypically to sugars on the cell surface Lectins are the third group of CAMs. They bind weakly and heterotypically to oligosaccharides of many types through the large extracellular domain. Selectins are lectins that are expressed in endothelial cells. Glycosyltransferases are lectin-related CAMs. They are enzymes that transfer monosaccharides to an oligo saccharide chain on an adjacent cell. In the absence of monosaccharides, the enzyme links one cell to another by binding to the oligosaccharide chain (can’t let go). In the presence of mono saccharides, the binding is lost. A simple way to regulate cell adhesion by mono saccharides.
Substrate adhesion molecules (SAMs) and the extracellular matrix (ECM) Spaces between cells are filled with ECM that consists of: 1. Amorphous ground substance: a gel-like material that absorbs water. 2. Meshwork of fibers that reinforce the ground substance. The ECM influences cell migration, cell shape, cell gene expression, and cell differentiation. Mesenchymal cells are surrounded by a diffuse ECM.Epithelial cells rest on a dense sheet of ECM called the basement membrane. The ECM is actively secreted by the cells living there. What molecules compose the ECM?
Glycosaminoglycans and proteoglycans form the amorphous ground substance Glycosaminoglycan: long unbranched polysaccharide chains composed of repeating units of disaccharides. One sugar is an amino sugar (n-acetyl glucosamine) and the other is a uronic acid (glucuronic acid). The most abundant glycosaminoglycans are hyaluronic acid, chondroitin sulfate, heparin, and heparin sulfate. Proteoglycans: glycosaminoglycans are covalently linked to core proteins The core proteins have have many side chains of glycosaminoglycans. They attract Na+ and water and expand to form gels that occupy space between cells. They also bind and selectively release growth factors. Glycosaminoglycan (hyaluronic acid) Proteoglycan
Fibrous glycoproteins make up the meshwork of the ECM Proteoglycansare carbohydrate with some protein (>50% carbohydrate). Glycoproteinsare proteins with some carbohydrate attached (> 50% protein). ECM consists of 3 major fibrous glycoproteins: Collagen: the most abundant protein in mammals (>25% total protein). There are numerous genes that encode different collagen molecules. Collagen forms very strong fibers that are abundant in bones, skin, and connective tissue. Fibronectin: a fibrous protein that has binding sites for cells and other ECM proteins. It links cells to the ECM. The RGD sequence of fibronectin (argenine, glycine, aspartate) binds to cells avidly. Fibronectin is important for motility. Laminins:abundant in basement membranes where they promote adhesion to many types of cells. How do cells stick to fibrous proteins of the ECM?
Integrins mediate adhesion to ECM Integrins are a family of transmembrane glycoproteins that are composed of 2 chains, a and b. There are 40 different types of a chains and 8 types of b chains that can combine to form a large number of different integrin molecules. The a chain has binding sites for Ca++ and Mg++ which are needed for integrins to adhere. The 2 subunits form the site that binds to the RGD domain on ECM. The cytoplasmic tail of integrins is connected to a linker protein that connects to the cytoskeleton. A bridge from ECM to cytoskeleton.
Cell surface proteoglycans also link cells to ECM These molecules often have an extracellular and intracellular domain. Proteoglycans consist of glycosaminoglycans linked to a core protein in the extracellular domain. These interact with collagen, fibronectin, and other ECM molecules. Proteoglycans can be released from the cell by cleavage at protease sensitive sites. Proteoglycans and integrins have another important function: They are receptors that transduce signals from the extracellular matrix to the nucleus. Syndecan is a common proteoglycan
If CAMs and SAMs were important for gastrulation, their expression might reflect that fact. 1. They might be expressed selectively on specific cells of the blastula that are destined to migrate to selected areas to form organs. 2. Expression might change as cells left the surface and ingressed or migrated into the blastocoel. 3. Blocking expression of CAMs or SAMs might interfere with the normal process of gastrulation.
CAM expression during gastrulation is correlated with cell fate Fate map: it is possible to predict which parts of the blastula will develop into specific structures after gastrulation. Expression map of CAMs: it is possible to localize expression of CAMs using in situ hybridization and immunostaining of the blastula. Cells with different fates express different CAMs. Cells destined to become neural tissue express high levels of N-CAM. Cells destined for epidermis express E-cadherin. The respective cell adhesion molecules are expressed before the cells actively start to form the adult tissue. This suggests that CAM expression is important in fate determination. During gastrulation, cells go where their CAMs lead them
If CAMs and SAMs were important for gastrulation, their expression might reflect that fact. 1. They might be expressed selectively on specific cells of the blastula that are destined to migrate to selected areas to form organs. 2. Expression might change as cells left the surface and ingressed or migrated into the blastocoel (changes in CAM drive movement). 3. Blocking expression of CAMs or SAMs might interfere with the normal process of gastrulation.
Changes in cell adhesion are important for gastrulation Gastrulation in the sea urchin is initiated by specific changes in cell adhesion. One of the first steps is ingression of mesenchymal cells from the vegetal plate into the blastocoel to form the skeleton of spicules. The mesenchymal cells lose their adhesion to hyaline and the adjacent nonmesenchymal blastomeres. They start to increase adhesion to the basement membrane and material within the blastocoel. These changes can be measured by isolating specific cells and testing adhesion in culture. E-cadherin is lost from the ingressing cells due to endocytosis of specific areas where it was expressed. Levels of b-catenin are also reduced on these cells.
CAMs promote formation of cell junctions CAMs allow cells to attach quickly but not tightly. They allow reversible changes in adhesion that aid in migration and intercalation. Cell junctions (tight junctions, desmosomes, gap junctions) are long term, tight, cell-cell attachments. They take longer to form. CAMs facilitate formation of cell junctions by ‘holding cells in place while the glue sets’. Compaction: cells of the blastula become polarized and form tight junctions which compacts the surface. Just before compaction, cells express high levels of E-cadherin where blastomeres touch (future tight junction). If blastulas are placed in medium with antibodies to E-cadherin to disrupt function, no compaction occurs. Mature tight junctions are associated with an area of abundant E-cadherin expression under the cell surface = the zonula adherens (think of a zipper).
If CAMs and SAMs were important for gastrulation, their expression might reflect that fact. 1. They might be expressed selectively on specific cells of the blastula that are destined to migrate to selected areas to form organs. 2. Expression might change as cells left the surface and ingressed or migrated into the blastocoel. 3. Blocking expression of CAMs or SAMs might interfere with the normal process of gastrulation.
Fibrous ECM components provide contact guidance to migrating cells during gastrulation The movement of cells during gastrulation may also depend upon expression of ECM. ECM allows migrating cells to attach transiently while moving over the surface. During gastrulation in amphibians, cells move into the blastocoel and migrate over the inside of the roof. If a portion of the roof is cut out and inverted, no movement of gastrulating cells occurs here. This suggests that some CAMs or SAMs may be missing. What molecules would this be?
Fibronectin on the inner roof of the blastocoel is critical for gastrulation Immunostaining of the blastocoel shows that fibronectin was expressed in abundance on the inner roof. Fibronectin binds to integrins on the membrane. Neutralizing antibody to fibronectin was injected into the blastocoel to test the role of fibronectin. This aborted gastrulation. Since no epidermal cells could migrate into the blastopore, many cells accumulated on the surface, forming deep folds. If an unrelated antibody was injected, there was no inhibition. Fibronectin binds integrins through an RGD sequence. Similar results were obtained by injecting the tripeptide RGD. Furthermore, blocking the integrin receptor with injected antibodies also inhibited gastrulation. Fibronectin is important for contact guidance of migrating cells during gastrulation.
CAMs and SAMs are major regulators of cell movement during gastrulation / morphogenesis. Do these molecules also directly influence gene expression or cell differentiation during morphogenesis? • During gastrulation cells migrate and are rearranged in the developing • embryo. Many new cell-cell contacts are established when cells reach their • new positions. • Do specific CAMs on one cell influence how the adjacent cell expresses • genes or undergoes differentiation? • 2. Would cells have different gene expression or differentiation depending upon the different types of ECM that they rested upon?
Neural differentiation is triggered by N-CAMs or N-cadherins N-cadherins and N-CAMs are expressed on presumptive neural tissue during and after gastrulation. What would happen if N-cadherin or N-CAM expression were experimentally abolished?
Neural differentiation is triggered by N-CAMs or N-cadherins N-cadherins and N-CAMs are expressed on presumptive neural tissue during and after gastrulation. What would happen if N-cadherin or N-CAM expression were experimentally abolished? Dominant negative mutants of N-cadherin: a mutant that blocks normal function dominantly (an N-cadherin molecule with the binding site for other cadherins cut off). This mutant was injected into a blastomere of presumptive neural tissue before gastrulation. The portion of the brain formed by daughter cells of this blastomere failed to develop. The other half of the brain that developed from uninjected blastomeres was normal. N-cadherin is critical for early brain development. Does N-cadherin directly influence neural differentiation?
PC12 cells resemble chromaffin cells which can differentiate into neurons. When they convert to the neural phenotype they express N-CAM and N-cadherin on their cell surface. When PC12 cells are grown on cells that do not express N-CAM or N-cadherin (3T3 cells) they retain the undifferentiated chromaffin phenotype. If the PC12 cells are grown on the same cells that have been transfected with N-CAM or N-cadherin genes, they convert to the neuronal phenotype. They form long dendrites and express neuronal genes. Differentiation is accompanied by opening of calcium channels
CAMs are necessary for neuronal differentiation Conversion of PC12 cells to the neuronal phenotype can be inhibited by adding antibodies that neutralize either N-CAM or N-cadherin on the 3T3 cells. Thus, adhesion through CAMs is necessary for neuronal differentiation in these cells. Neuronal differentiation is also inhibited by drugs that block calcium channels. This suggests that intracellular signaling by calcium is an important requirement for neuronal differentiation.
CAMs and SAMs are major regulators of cell movement during gastrulation / morphogenesis. Do these molecules also directly influence gene expression or cell differentiation during morphogenesis? • During gastrulation cells migrate and are rearranged in the developing • embryo. Many new cell-cell contacts are established when cells reach their • new positions. • Do specific CAMs on one cell influence how the adjacent cell expresses • genes or undergoes differentiation? • 2. Would cells have different gene expression or differentiation depending upon the different types of ECM that they rested upon?
Collagen directly activates epithelial cells to form stroma During development, the cornea (outer eye) and lens (inner eye) interact. The cornea consists of outer epithelial cells and inner stromal cells. Formation of the inner layer depends upon interaction with the underlying lens which is covered with collagen. This interaction has been studied in culture. When corneal cells are grown on top of the lens (i.e., in contact with collagen), they differentiate to form a lower layer of stroma. When corneal cells are cultured on artificial substrate, no stroma forms. When the cells are cultured on a collagen substrate, a normal stroma develops. Interaction with collagen is critical for corneal differentiation.