Download

1 / 31
310 likes | 484 Views
Exploring the Universe of Protein Structures beyond the Protein Data Bank. Flavio Seno Dipartimento di Fisica e Astronomia Universita’ di Padova. Means, Methods and Results in the Statistical Mechanics of Polymeric Systems. Toronto 21-22 June 2012.

E N D
Exploring the Universe of Protein Structures beyond the Protein Data Bank Flavio Seno Dipartimento di Fisica e Astronomia Universita’ di Padova Means, Methods and Results in the Statistical Mechanics of Polymeric Systems. Toronto 21-22 June 2012
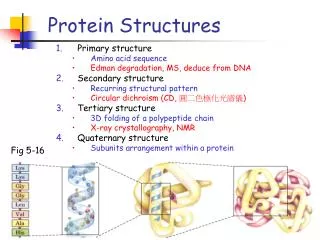
What are their distinctive properties?secondary structuresstabilized by hydrogen bondsFolds:arrangements of secondary structures in the spaceThere is a limited set of folds:same folds used to perform different functionsThere is not macroscopic evolution:multiple separate discoveries during the course of evolution Protein structures
~ 7000 structures (new sequences) determined every year Platonic folds : intrinsic features of the order of nature (Denton and Marshall ,Nature 2002)
“SIMILARITY OF PROTEIN STRUCTURES IMPOSED BY SOME PHYSICAL REGULARITIES” (Finkelstein-Ptitsyn 2002) Are protein folds determined only by physical and geometrical laws ( crystal structures) and not by the chemistry of the amino-acid sequence? Is it possible to reproduce them in terms of general principles? Maybe through an homopolymer that captures the main common features of all the aminoacids? Are the observed folds in a one to one correspondence with the whole possible fold universe? If not, why? Is there a selection principle?
- C Representa tion a Minimal Coarse-Grained ModelT.X. Hoang, L. Marsella, A. Trovato, J.R. Banavar, A. Maritan, F.S. PNAS, vol 103, 6883 (2006) • Excluded volume (self-avoiding tube) • Hydrogen bonding geometric constraint • Hydrophobic interaction • Local bending penalty
Homopolypeptide structures in the ‘marginally compact’ phase (compact + h-bonds) are protein-like Ground State Phase Diagram
METADYNAMICS A Laio, M Parrinello, Escaping free-energy minimaPROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF, 99, 12562 (2002) HOW TO FIND STABLE MINIMA WHICH ARE SEPARATED BY BARRIERS THAT CANNOT CLEARED IN THE AVAILABLE SIMULATION TIME THE METHOD IS BASED ON AN ARTIFICIAL DYNAMICS (METADYNAMICS) 1) IDENTIFY COLLECTIVE VARIABLES S WHICH ARE ASSUMED TO PROVIDE A RELEVANT COARSE GRAINED DESCRIPTION OF THE SYSTEM 2) TO BIAS THE DYNAMICS ALONG THESE VARIABLES. S 3) RUN IN PARALLEL SEVERAL MOLECULAR DYNAMICS EACH BIASED WITH A METADYNAMIC POTENTIAL 4) SWAPS OF THE CONFIGURATIONS
ATOMISTIC MODEL 60 AMINO ACIDS POLYVALINE (VAL60) • Why VAL? (is small but not too much) • MD simulations with AMBER force field • and package GROMACS • Bias-exchange METADYNAMICS with 6 • replicas • Six collective variables linked to secondary • structure elements
50 microseconds molecular dynamics simulationWe generate an ensemble of 30000 all-atom conformations SIGNIFICANT SECONDARY STRUCTURE CONTENT AND SMALL RADIUS OF GYRATION
We verify they are local minima also for ALA-60Structural quality resembles that of real protein QUALITY MEASURE G-FACTOR RAMACHANDRAN PLOT FRAGMENT DISTANCE < 0.6 A H-BOND ENERGY COMPUTED WITH PROCHECK 0.7 A
FIRST RESULT FINDING BY MOLECULAR DYNAMICS AT AN ALL-ATOM LEVEL A LIBRARY OF 30000 PROTEIN LIKE STRUCTURES http://datadryad.org/handle/10255/dryad.1922
RELATION BETWEEN VAL60 AND REAL PROTEINS The Class Architecture Topology and Homologous superfamily protein structure classification (CATH) is one of the main databases providing hierarchical classification of protein domain structures. 300 FOLDS 40 < L<75 SIMILARITY: TM-SCORE (Zhang Scolnick 2005) ALLIGNMENTS OF SECONDARY STRUCTURES ALLOWING INSERTIONS AND DELETIONS (COVERAGE) TM=0.45 MINIMIZATION OF THE RELATIVE DISTANCE BETWEEN ALIGNED RESIDUES (RMSD)
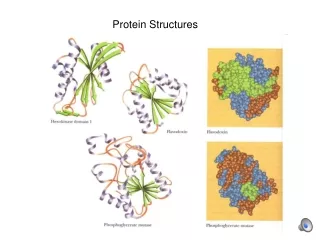
1x9b 1ib8 1g29 COMPARISON VAL60 VS CATH 40 < L < 75 300 FOLDS 1uxy
SECOND RESULT THE COMPUTATIONAL SETUP USED IN THIS WORK ALLOW US TO EXPLORE THE MAJORITY OF THE FOLDS IN NATURE (AT LEAST FOR THESE LENGTHS)
COMPARISON POLYVAL VS CATH NOT ALL VAL60 ARE PRESENT IN CATH!!!!!!! TM =0.45 VAL60 7000 CATH 300
THIS MIGHT JUST DEPEND ON THE CHOSEN SIMILARITY THRESHOLD DO STRUCUTRAL DESCRIPTORS DISCRIMINATE BETWEEN CATH AND VAL60? CONTACT ORDER: Average sequence separation between contacting residues (related to folding rates Plaxco Simons Baker 1998) • Real protein strucures were selected under a bias towards low CO • - protein structures are selected to be topologically less entangled
THIRD RESULT THERE IS NO ONE-TO-ONE CORRESPONDENCE BETWEEN PDB LIBRARY AND THE ENSEMBLE OF COMPACT STRUCUTRES WITH SIGNIFICANT SECONDARY STRUCUTURE CONTENT (VAL60)
SUMMARY • VAL60 SET IS REPRESENTATIVE OF REAL PROTEINS (PROTEINS FOLDS SELECTED BY GEOMETRY AND SIMMETRY AND NOT BY CHEMISTRY OF THE SEQUENCE) • KNOWN FOLDS FORM ONLY A SMALL FRACTION OF THE FULL DATABASE • NATURAL FOLDS ARE CHARACTERIZED BY SMALL CONTACT ORDER WHY KINETIC ACCESSIBILITY HIGHER CO HIGHER TENDENCY TO AGGREGATE?
APPLICATIONS • REALISTIC DECOYS • DESIGN NEW PROTEINS • CHECK PREDICTIONS IN SYNTHETIC BIOLOGY • MODELS FOR MISFOLDED STRUCTURES RELATED TO NEURODEGENERATIVE DISEASES
COLLABORATORS • PILAR COSSIO (NIH WASHINGTON) • ALESSANDRO LAIO (SISSA TRIESTE) • DANIELE GRANATA (SISSA TRIESTE) • FABIO PIETRUCCI (CECAM – LAUSANNE) • AMOS MARITAN (PADOVA) • ANTONIO TROVATO (PADOVA) Plos Computational Biology vol.6 e 1000957 (2010) Scientific Reports 2, Art. No. 351 (2012)
CORRELATION BETWEEN POTENTIAL ENERGY AND CONTACT ORDER FOR VAL60 AND ALA60 STRUCUTRES
Distribution of the radius of gyration for the VAL60, VAL60+WATER,ALA60 and CATH 55–65 sets of structures.
Cα RMSD distributions for the 30,000 VAL60 and the 1500 ALA60 minimized through SD.
Probability of finding a structure in the VAL60 trajectory for different CO classes.
Bias Exchange Metadynamics S Piana, A Laio, A bias-exchange approach to protein folding JOURNAL OF PHYSICAL CHEMISTRY B, 111, 4553 (2007) IT IS AN APPROACH DESIGNED FOR ACCELATING RARE EVENTS IN VERY COMPLEXES CASES IN WHICH THE VARIABLES THAT ARE RELVANT FOR THE PROCESS ARE MORE THAN 2 OR 3 • List all the collective variables • Run in parallel several molecular dynamics each biased with a metadynamic potential • Swaps of the configuration
Are compact hydrogen-bonded polypetide structures in one-to-one correspondence with protein structuresfrom the Protein Data Bank (PDB)? Homopolypeptide ( side chain:C-beta atoms) with a very minimal potential consisting of H-bonding, excluded volume, and a uniform, pairwise attractive potential between side chains. YES!? PNAS 103, 2605-2010 (2006)