Download

1 / 105
1.06k likes | 1.22k Views
D-Metabolic Pathways. Do not rely on passive reading and highlighting/underlining of the textbook. Do not sit and stare at the handouts Do not try to read 50 review books. (Make your own review book instead!) Do focus on identifying key concepts

E N D
Do not rely on passive reading and highlighting/underlining of the textbook. • Do not sit and stare at the handouts • Do not try to read 50 review books. (Make your own review book instead!) • Do focus on identifying key concepts • Do actively draw and redraw pathways and connections • Do learn to identify relevant information
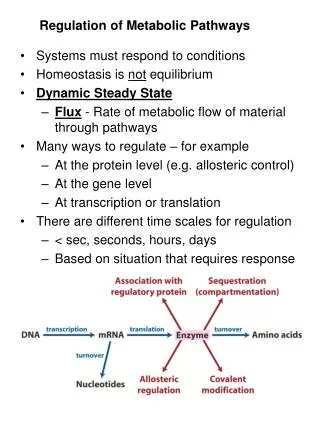
Do prioritize: • What is the purpose of a pathway? • What are the starting and ending molecules? • Where is the pathway (in the cell, in a tissue, in an organ system)? • How does the pathway connect to other pathways? • What metabolic conditions turn the pathway on and off? • What are the control points for regulating the pathway? • reactants, products and enzyme name of each regulatory step • additional regulatory molecules involved (vitamins, cofactors) • make sure you know every step that makes or uses ATP • What structural features are important for the function and interaction of specific regulatory molecules in a pathway? • What biochemical techniques are used to study these pathways? • What specific drugs or diseases associated with the pathway?
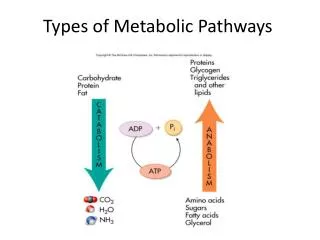
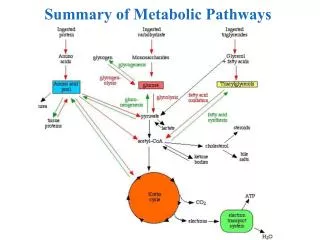
METABOLIC PATHWAYS • Glycolysis • Gluconeogenesis • Citric Acid Cycle (Krebs Cycle) • Glycogen Metabolism • Hexose Interconversions • Electron Transport Chain • Oxidative Phosphorylation • Pentose-Phosphate Shunt
GLYCOLYSIS • Oxidation of glucose is known as Glycolysis. • Either AerobicàPyruvate • Anaerobicà Lactic Acid • Occurs in the Cytosol • Overall Rxn: • Glucose + 2 ADP + 2 NAD+ + 2 Pi à 2 Pyruvate + 2 ATP + 2 NADH + 2 H+ • NADH generated during glycolysis is used to fuel mitochondrial ATP synthesis via oxidative phosphorylation. Does not pass through mitochondrial membrane • 2 ATP generatedàglycerol phosphate shuttle • 3 ATP generatedàmalate-aspartate shuttle • If used to transport the electrons from cytoplasm NADH into the mitochondria.
Key Reactions • Hexokinase • Found in the cytosol of most tissues • Low specificity: it’s a “hoe” for hexoses • Low Km: high affinity for glucose • Inhibited by Glucose-6-phosphate • Glucokinase: Found in the Liver and pancreatic b cells • Also a ‘hexokinase’ • High specificity for glucose • High Km • inhibited by fructose-6-phosphate
Regulation of Glycolysis • Hexokinase, PFK-1 and PK all proceed with a relatively large free energy decrease. These non-equilibrium reactions of glycolysis would be ideal candidates for regulation of the flux through glycolysis. • Hexokinase is not key because of G6P is generated by glycogenolysis • PK reaction is reversed in Gluconeogenesis • Therefore rate limiting step in glycolysis is the reaction catalyzed by PFK-1. • PFK-1 is a tetrameric enzyme that exist in two conformational states termed R and T that are in equilibrium. • ATP is both a substrate and an allosteric inhibitor of PFK-1. F6P is the other substrate for PFK-1 and it also binds preferentially to the R state enzyme. ATP binds the T state.
The inhibition of PFK-1 by ATP is overcome by AMP which binds to the R state of the enzyme and, therefore, stabilizes the conformation of the enzyme capable of binding F6P. • The most important allosteric regulator of both glycolysis and gluconeogenesis is fructose 2,6-bisphosphate, F2,6BP, which is not an intermediate in glycolysis or in gluconeogenesis. • Also important to note that Insulin/Glucagon ratio i.e. fed/starve state, regulate Pyruvate Kinase activity. The last enzyme in the pathway. • Glucagon: high in starvation, b/cos blood glucose levels are low, therefore it favors gluconeogenesis in Liver. • Insulin: on the contrary favors glycolysis.
Glycolysis • Key points about the Shuttle System: • Malate-Asparate shuttle is the primary system • By default Glycerol shuttle is secondary • Two enzymes are involved in this shuttle: • 1.cytosolic version of the enzyme glycerol-3-phosphate dehydrogenase (glycerol-3-PDH) which has as one substrate, NADH. • 2.mitochondrial form of the enzyme which has as one of its' substrates, FAD+. Since the electrons from mitochondrial FADH2 feed into the oxidative phosphorylation pathway at coenzyme Q (as opposed to NADH-ubiquinone oxidoreductase [complex I]) only 2 moles of ATP will be generated from glycolysis. G3PDH is glyceraldehyde-3-phoshate dehydrogenase.
Malate -Asp Shuttle • The electrons are "carried" into the mitochondria in the form of malate. Cytoplasmic malate dehydrogenase (MDH) reduces oxaloacetate (OAA) to malate while oxidizing NADH to NAD+ • Cytoplasmic malate dehydrogenase (MDH) reduces oxaloacetate (OAA) to malate while oxidizing NADH to NAD+. • Malate then enters the mitochondria where the reverse reaction is carried out by mitochondrial MDH • mitochondrial OAA goes to the cytoplasm to maintain this cycle ; must be transaminated to aspartate (Asp) with the amino group being donated by glutamate (Glu). The Asp then leaves the mitochodria and enters the cytoplasm. The deamination of glutamate generates a-ketoglutarate (a-KG) which leaves the mitochondria for the cytoplasm. • When the energy level of the cell rises, the rate of mitochondrial oxidation of NADH to NAD+ declines and therefore, the shuttle slows.
The synthesis of F2,6BP is catalyzed by the bifunctional enzyme phosphofructokinase-2/fructose-2,6-bisphosphatase (PFK-2/F-2,6-BPase). • In the nonphosphorylated form the enzyme is known as PFK-2 and serves to catalyze the synthesis of F2,6BP by phosphorylating fructose 6-phosphate. • The result is that the activity of PFK-1 is greatly stimulated and the activity of F-1,6-BPase is greatly inhibited. More glycolysis! • When the bifunctional enzyme is phosphorylated it no longer exhibits kinase activity, but a new active site hydrolyzes F2,6BP to F6P and inorganic phosphate. • This enzyme is regulated by ProteinKinase A, which is a cyclic AMP dependent enzyme. cAMP is generated depending on the hormonal changes in the body. Eg. With Glucagon, high cAMP thus PKA is active thus less glycolysis. • In addition to these Pyruvate Kinase is activated by F1,6BP and inhibited by ATP.
Substrates for Gluconeogenesis: Lactate, pyruvate, glycerol, propionny-CoA and certain Amino Acids but never FAT!!! • The Cori cycle involves the utilization of lactate, produced by glycolysis in non-hepatic tissues, (such as muscle and erythrocytes) as a carbon source for hepatic gluconeogenesis. In this way the liver can convert the anaerobic byproduct of glycolysis, lactate, back into more glucose for reuse by non-hepatic tissues. Note that the gluconeogenic leg of the cycle (on its own) is a net consumer of energy, costing the body 4 moles of ATP more than are produced during glycolysis. Therefore, the cycle cannot be sustained indefinitely.
The glucose-alanine cycle is used primarily as a mechanism for skeletal muscle to eliminate nitrogen while replenishing its energy supply. Glucose oxidation produces pyruvate which can undergo transamination to alanine. This reaction is catalyzed by glutamate-pyruvate transaminase, GPT (also called alanine transaminase, ALT in Figure).
Regulation of Gluconeogenesis • See regulation of Glycolysis via F2,6 P • Do not forget Hormonal regulations: Insulin and Glucagon • Other things to keep in mind • Pyruvate carboxylase is present in mitochondria, requires Biotin as a cofactor to convert Pyruvateà OAA • MDH present in mitochondria, OAA to malate, then MDH present in cytosol converts malate back to OAA • OAA is then converted to PEP, as shown in the previous slide. • Pyruvate Carboxylase: inhibited by ADP and activated Acetyl CoA • PEP Carboxykinase in the cytosol is inhibited by ADP
TCA /Citric Acid/KREBS Cycle • The cycle is located in the mitochondria • All cells have a mitochondria except RBCs • This is the Final common pathway of oxidative metabolism • Acetyl coenzyme A condenses with OAA to begin the cycle. Catabolism of CHO, Fats and Proteins provide the acetyl CoA • The bulk of ATP used by many cells to maintain homeostasis is produced by the oxidation of pyruvate in the TCA cycle • During this oxidation process, reduced NADH and reduced FADH2 are generated. The NADH and FADH2 are principally used to drive the processes of oxidative phosphorylation, which are responsible for converting the reducing potential of NADH and FADH2 to the high energy phosphate in ATP
The PDH complex requires 5 different coenzymes: CoA, NAD+, FAD+, lipoic acid and thiamine pyrophosphate (TPP) . Three of the coenzymes of the complex are tightly bound to enzymes of the complex (TPP, lipoic acid and FAD+) and two are employed as carriers of the products of PDH complex activity (CoA and NAD+).pyruvate + CoA + NAD+ à CO2 + acetyl-CoA + NADH + H+
The TCA cycle showing enzymes, substrates and products. The abbreviated enzymes are: IDH = isocitrate dehydrogenase and a-KGDH = a-ketoglutarate dehydrogenase. The GTP generated during the succinate thiokinase (succinyl-CoA synthetase) reaction is equivalent to a mole of ATP by virtue of the presence of nucleoside diphosphokinase. The 3 moles of NADH and 1 mole of FADH2 generated during each round of the cycle feed into the oxidative phosphorylation pathway. Each mole of NADH leads to 3 moles of ATP and each mole of FADH2 leads to 2 moles of ATP.
Overall Stoichiometry of TCA • acetyl-CoA + 3NAD+ + FAD + GDP + Pi + 2H2O ----> 2CO2 + 3NADH + FADH2 + GTP + 2H+ + HSCoA • The GTP generated by Succinyl CoA SYNTHETASE IS VIA SUBSTRATE LEVEL PHOSPORYLATION. • Regulation of TCA:Regulation of the TCA cycle like that of glycolysis, occurs at both the level of entry of substrates into the cycle as well as at the key reactions of the cycle. Fuel enters the TCA cycle primarily as acetyl-CoA. The generation of acetyl-CoA from carbohydrates is a major control point of the cycle. This is the reaction catalyzed by the PDH complex • PDH complex is inhibited by acetyl-CoA, ATP, and NADH • PDH activated by non-acetylated CoA (CoASH) and NAD+. • The pyruvate dehydrogenase activities of the PDH complex are regulated by their state of phosphorylation. This modification is carried out by a specific kinase (PDH kinase) and the phosphates are removed by a specific phosphatase (PDH phosphatase). • The phosphorylation of PDH inhibits its activity which leads to decreased oxidation of pyruvate. • PDH kinase is activated by NADH and acetyl-CoA and inhibited by pyruvate, ADP, CoASH, Ca2+ and Mg2+. The PDH phosphatase, in contrast, is activated by Mg2+ and Ca2+ • Citrate Synthase: inhibited by ATP and citrate • Isocitrate Dehydrogenase: Isocitrate, AMP, ADP activates, ATP and NADH inhibits • A-ketoglutarate dehydrogenase: succinoyl CoA and NADH inhibits • CindyIsKinkySoSheFornicatesMoreOften
ELECTRON TRANSPORT AND OXIDATIVE PHOSPHORYLATION • Each turn of TCA cycle generates 3NADH and 1 FADH2 • Electron transport and oxophos occurs in the mitochondria • NADH and FADH2 ultimately pass electrons to O2 and produce H2O. • NADH + (1/2)O2 + H+ -->NAD+ + H2O ~ -52.6kcal/mol • ADP + PàATP ~ +7.3kcal/mol • Energy from NADH can be used to drive synthesis of ATP several times. • Important again to remember this is an oxidation-reduction reaction thus our friend Nerst is back: • DeltaG' = -nFDE'
Electron Transport is coupled to Oxidative Phosphorylation • The idea of coupling is explained by Mitchell’s CHEMIOSMOTIC HYPOTHESIS • Basically coupling electron flow through the ETC to ATP synthesis • The Respiratory complexes are proton pumps. As electrons pass through complexes I, III, and IV, hydrogen ions are pumped across the inner mitochondrial membrane into the intermembrane space. • The proton concentration in the intermembrane space increases relative to the mitochondrial matrix • This generates a proton-motive force as a result of 2 factors: 1) Difference in pH and 2) Difference in electrical potential, delta si, between intermembrane space and the mitochondrial matrix. • ATP synthetase complex (complex V): Hydrogen ions pass back into the matrix through V, this drives ATP synthesis. • NADHà 3ATP • FADH2à 2 ATP: note bypass of Complex 1 • ATP synthesized in the matrix is transported out of the matrix via an ATP/ADP translocase (an antiport) also coupled to proton motive force.
Inhibitors of Oxidative Phosphorylation • Rotenone: e- transport inhibitor Complex I • Amytal: e- transport inhibitor Complex I • Antimycin: A e- transport inhibitor Complex III • Cyanide: e- transport inhibitor Complex IV • Carbon Monoxide: e- transport inhibitor Complex IV • Azide e- transport inhibitor Complex IV • 2,4,-dinitrophenol: Uncoupling agent transmembrane H+ carrier • Pentachlorophenol: Uncoupling agent transmembrane H+ carrier • Oligomycin: Inhibits ATP synthase • Thermogenin: also an uncoupler, component of brown fat • Malonate inhibits Complex II • There are others in your handout take a look at them.
SOME MORE STUFF • TCA cycle is regulated by the ratio of ADP, Pi/ ATP • Under resting conditions, with a high cell energy charge, the demand for new synthesis of ATP is limited and, although the Proton Motive Force is high, flow of protons back into the mitochondria through ATP synthetase is minimal. When energy demands are increased, such as during vigorous muscle activity, cytosolic ADP rises and is exchanged with intramitochondrial ATP via the transmembrane adenine nucleotide carrier ADP/ATP translocase. Increased intramitochondrial concentrations of ADP cause the Proton Motive Force to become discharged as protons pour through ATP synthetase, regenerating the ATP pool. • The rate of electron transport is dependent on the PMF • ANY BLOCKADE AT ANY POINT IN THE ELECTRON TRANSPORT CHAIN STOPS ATP SYNTHESIS!!!!!!!!!
Choose the INCORRECT statement concerning the ATP-ADP cycle and the study of bioenergetics in the human body: • a. One half of the ATP-ADP cycle involves the coupling the energy derived from the hydrolysis of the high energy phosphate bonds of ATP to endergonic reactions so that they will occur spontaneously. • b. The work that requires energy derived from ATP hydrolysis includes the transport of electrons down the electron transport chain. • c. One half of the ATP-ADP cycle involves the generation of ATP that starts with the formation of reduced coenzymes like NADH and FADH2and the ultimate transfer of their electrons to oxygen • d. An important part of oxidative phosphorylation and ATP biosynthesis is the generation of an electrochemical gradient across the inner membrane of the mitochondria. • Many catabolic reactions, like the TCA cycle and fatty acid oxidation, provide the reduced coenzymes for the start of oxidative phosphorylation and ATP biosynthesis
Since electron transport and oxidative phosphorylation are tightly coupled, which one of the following mechanisms BEST explains how ADP regulates the rate of electron transport during oxidative phosphorylation? • a. AMP concentrations are increased as ADP concentrations fall • b. Low [ADP] accelerates the Krebs (TCA) cycle reaction rates, thereby providing more NADH to activate electron transport • c. The transmembrane proton gradient is dissipated with low [ADP] • d. The ATP/ADP antiport system is not functional when mitochondrial [ADP] is low • e. Proton translocation across the inner mitochondrial membrane is decreased when ATP-synthase lacks bound ADP and Pi, secondarily retarding electron transport
You isolate mitochondria from a group of patients that present with lactic acidosis and muscle weakness, and show that they are unable to: (1) oxidize reduced coenzyme Q, (2) translocate protons across their mitochondrial membranes to the intennembrane space against a concentration gradient with succinate added as the substrate, and (3) reduce cytochrome c. The biochemical defect in these patients most likely resides in their ... ? • A. Complex I (NADH dehydrogenase) • B. Complex II (succinate-Q reductase) • C. Complex III (cytochrome b-c1) • D. Complex IV (cytochrome oxidase) • E. Complex V (F1F0 ATPase)
Which of the following orderings #1 - #5 of the various components of the electron transport chain and oxidative phosphorylation will effectively allow the development of an electrochemical potential sufficient to drive the generation of high energy phosphate bonds between ADP and Pi? • 1. FMN, NADH dehydrogenase, ubiquinone, cytochrome c, cytochrome oxidase, F1F0-ATPase • 2. Complex I, Complex III, ubiquinone, cytochrome a1-a3, cytochrome c, Complex IV, Complex V • 3. FAD(2H)/succinate dehydrogenase, Coenzyme Q, cytochrome b-cl, cytochrome c, cytochrome a1-a3, F1F0-ATPase • 4. NADH dehydrogenase, CoQ, cytochrome b-cl, cytochrome c, cytochrome oxidase, ATP synthase • 5, NADH dehydrogenase, CoQ, cytochrome c, cytochrome oxidase, cytochrome b-cl, F1F0-ATPase • a. Both #1 and #2 • b. Both #3 and #4 • c. Only #4 • d. Only #3 • e. None of the above
As a skilled cell biologist and biochemist, you cleverly devise a method for experimentally separating the F1 portion of ATP synthase from the membrane-bound Fo fragment in intact mitochondria. Which of the following metabolic effects do you observe? • a. Electron transport and oxygen consumption are inhibited • b. Electron transport and phosphorylation of ADP remain tightly coupled • c. The inner mitochondrial membrane remains impermeable to protons • d. Protons pass through the membrane-bound Fo fragment, but they do not sustain any ATP formation • e. The F1 fragment forms ATP at an accelerated rate until ADP is depleted or the proton gradient is dissipated
Which of the following groups of enzymatic reactions, enzymes and substrates comprise important anaplerotic pathways for 4-carbon intermediates critical to the citric acid (TCA) cycle in the liver, muscle and nervous tissues? • a. conversion of pyruvate to acetyl CoA via pyruvate dehydrogenase and glutamate to a-ketoglutarate via transaminases • b. conversion of cc-ketoglutarate to glutamate and GABA • c. production of ketone bodies (acetoacetate and P-hydroxybutyrate) • d. conversion of pyruvate to oxaloacetate via pyruvate carboxylase, biotin, bicarbonate ion, and ATP • e. both (A) and (D)
Regulation of tricarboxylic acid cycle activity in vivo may involve the concentration of all of the following EXCEPT: • acetyl CoA • ADP. • ATP. • CoA. • oxygen.
NAD+ can be regenerated in the cytoplasm if NADH reacts with any of the following EXCEPT: • pyruvate. • dihydroxyacetone phosphate. • oxaloacetate. • the flavin bound to NADH dehydrogenase. • phosphoglycerate kinase.
Glucokinase: • has a Km considerably greater than the normal blood glucose concentration.. • is found in muscle. • is inhibited by glucose 6-phosphate. • is also known as the GLUT-2 protein. • has glucose 6-phosphatase activity as well as kinase activity.
A 7yr old female presents with anxiety, dizziness, sweating and nausea following brief periods of exercise. The symptoms are relieved by eating and do not occur if the patient is frequently fed small meals. Blood analysis indicates she is hypoglycemic following brief period of fasting, alanine fails to increase blood sugar, fructose or glycerol administration restores glucose to normal? • What Pathway is affected, which enzyme could it be? How would you confirm your speculation?
After the BIOCHEM exam you and your friends decided to only drink “liquid-fire” (Bacardi 151) for the rest of the evening. The next morning you manage to wakeup with terrible ‘hangover’. Which of these molecules is most responsible for your hangover? • Lactic Acid • Pyruvate • Acetate • Acetyladehyde • Ethanol
ADH àalcohol dehydrogenase • AcDHà acetyladehyde dehydrogenase • Acetaldehyde forms adducts with Proteins, nucleic acids, and other compounds results in hangover. • NADH/NAD+ imbalance causes Liver to over work. • Diversion of gluconeogenesis by Lactic Acid dehydrogenase decreases ability of Liver to deliver glucose to the blood. • In addition, there is increased synthesis of FAT. Acetate + CoA gives you acetyl-CoA which is a precursor for Fatty acid sythesis. You already have enough NADH to go to work. So let the FATTYLIVER BEGIN! HepatoMEGALLY! Lets go!
CLINICAL CORRELATIONS • Riboflavin Deficiency • FMN and FAD are both synthesized from riboflavin, which contains the electron-accepting ring structure of FAD • Severe Riboflavin deficiency decreases the ability of mitochondria to generate ATP via oxidative phosphorylation • In general, impairment of Complex I (NADH Dehydrogenase) induces formation of mitochondria with structural abnormalities. • Iron Deficiency Anemia • Characterized by decreased levels of Hb and other heme containing proteins in blood. • Iron-containing cytochromes and Fe-S centers of ETC are decreased as well. • Fatigue partly due to impaired ETC for ATP generation • ETC inhibitors at specific sites • Rotenone and Amytal block Complex I • Antimycin blocks cytochrome b1 in Complex III • Cyanide blocks cytochrome a/a3 in Complex IV. Prevents reduction of e- from reduced cytochrome c. • CO binds to reduced iron of cytochrome oxidase • Cyanide Poisoning • CN- causes a rapid and extensive inhibition of ETC at the cytochrome oxidase step. Prevents O2 from serving as the final e- acceptor. • Mitochondrial respiration and energy production cease, resulting in cell death • Occurs from tissue asphyxiation, most notably in the Nervous System • Treatment: nitrites administered to convert oxyHb to MetheHb, which can then compete with cytochrome a,a3 for the CN-, forming a complex.
Oxidative Phosphorylation II “the uncoupling of ETC and Ox-Phos” • Uncoupling of ETC with Ox-Phos • Proton gradient from ETC coupled to ATP production from Oxidative Phosphorylation. If uncoupled and proton gradient dissipated, ATP and ADP concentrations lose their ability to regulate the rate of e- transport. • Uncouplers:proton ionophores, which rapidly transport H+ from cytosolic to matrix side of inner mitochondiral membrane • DNP – picks up H+ on cyto side, drops H+ on matrix side • Oligomycin: inhibits F1F0-ATPase…ATP synthesis stops. • Respiration and transport are blocked • Addition of an uncoupler (DNP) induces initiation of O2 consumption…ETC continues but w/o ATP synthesis since the pathways are uncoupled. • Brown Adipose Tissue and Thermogenesis • Large deposits of brown fat around vital organs (in human infants)…specialized for ‘non-shivering thermogenesis.’ • Cold or excessive food intake stimulates NE release • Then Thermogenin, proton conductance uncoupler, is activated, pumping H+ back into mitochondria…dissipating the gradient. • ETC is induced, increasing rate of NADH and FADH2 oxidation, which generates more heat = biological heating pad
Hyperthyroidism – Grave’s Disease • Thyroid hormone influences bioenergetics via actions on mitochondrial ox phos. • In Hyperthyroidism, energy derived from ox. Phos is significantly less than normal. • Thryoid causes ‘uncoupling’ of Ox Phos. • Results in increased heat production – patients complain of feeling hot and sweaty. • Salicylate (aspirin) poisoning • At high concentrations, salicylate can partially uncouple mitochondrial Ox Phos. • Decreased ATP [ ] and increased cytosolic AMP induce glycolysis • Results in increased blood pyruvate and lactate and metabolic acidosis and fever • Myoclonic Epileptic Ragged Red Fiber Disease (MERRF) • Debilitating, progressive spontaneous muscle jerking • Mitochondrial myopathy with enlarged, abnormal mitochondria • Neurosensory hearing loss, dementia, hypoventilation, mild cardiomyopathy • Maternal inheritance (sex linked) • Impaired energy metabolism….lactic acidosis
Pentose Phosphate Pathway • Hemolysis caused by Reactive Oxygen Species (ROS) • G6PD deficiency in pentose phosphate pathway • Causes increased production of radicals from GSH, since can’t produce sufficient NADPH to re-reduce glutathione….result in hemolysis • Heinz Bodies in RBCs • Due to G6PD deficiency • RBCs need the enzyme to re-reduce glutathione with NADPH to protect against oxidative stress • ROS peroxidation of membrane lipids lyses the RBC membrane • G6PD Mediterranean disease most severe G6PD deficiency • Lecture 21 – Monosaccharides and interconversion of sugars • Classical Galactosemia • Deficiency of Galactosyl-1-P uridylyltransferase • Accumulation of G-1-P in tissues and inhibition of glycogen metabolism, which require UDP-sugars • Higher level of galactose in blood and urine • More serious form • Non-Classical Galactosemia • Galactokinase deficiency • Unable to convert galactose to galactose-1-P
Glycogen Synthesis • Glucose Toxicity • Dysfunction of glycogen synthase • Due to hyperglycemia…produces insulin resistance • Due to production of hexosamines that inhibit hexokinase, protein phosphatase 1, and glycogen synthase. • Lecture 23 – Glycogen Degradation • Von Gierke’s Disease • Defective G-6-Phosphatase enzyme • Increased amount of glycogen, normal structure • Affects liver and kidney • Massive enlargement of the liver. Severe hypoglycemia, ketosis, hyperuricemia, hyperlipemia. • Lecture 24 – Glucose/Glycogen Regulation • Type I – Insulin-dependent diabetes mellitus • Hyperglycemic • Continuous glucagon expression causes ketogenesis, lipolysis, and gluconeogenesis. • Hyperchylomicronemia occurs (liver TG syn and VLDL transport faster than adipose LPL breakdown of TG) • Risk of ketoacidosis • Type II – Noninsulin-dependent Diabetes Mellitus • Hyperglycemic • Peripheral tissues insulin resistant • Glucose accumulates in blood due to poor uptake by peripheral tissues, particularly muscles • Hypertriacylglycerolemia, which results from increase of VLDL without hyperchylomicronemia. New FA and VLDL synthesized in liver instead of increased delivery of fatty acids from adipose tissue.
Pentose Phosphate Pathway What is the PPP and why is it important?
Pentose Phosphate Pathway What is the PPP and why is it important? The pentose phosphate pathway is primarily an anabolic pathway that utilizes the 6 carbons of glucose to generate 5 carbon sugars and reducing equivalents
Pentose Phosphate Pathway • To generate reducing equivalents, in the form of NADPH, for reductive biosynthesis reactions within cells • To provide the cell with ribose-5-phosphate (R5P) for the synthesis of the nucleotides and nucleic acids • Although not a significant function of the PPP, it can operate to metabolize dietary pentose sugars derived from the digestion of nucleic acids as well as to rearrange the carbon skeletons of dietary carbohydrates into glycolytic/gluconeogenic intermediates