Download

1 / 29
290 likes | 301 Views
The cell, plasma membrane, endoplasmic reticulum. prof. Dr. Pál Röhlich. ÁOK 2017/2018, Semester I: Introduction to cell biology 11.13. 14. 09. 2017. The cell. Living organisms consist of cells and their products.

E N D
The cell, plasma membrane, endoplasmic reticulum prof. Dr. Pál Röhlich ÁOK 2017/2018, Semester I: Introduction to cell biology 11.13. 14. 09. 2017
The cell Living organisms consist of cells and their products. The cell is the smallest structural and functional unit, which still shows the basic phenomena of life and is capable of individual life: Examples: it synthetizes macromolecules using a genetic program, takes up substances from the environment to obtain energy and to build up its own structure, the cell adapts itself to its varying environment, it carries out active movements, is able to reproduce itself through cell division and transfers its genetic program to daughter cells, registers signals from its surrounding and responds to them, …) Size measures: 1 mikrometer (μm) = 10-3 mm, 1 nanometer (nm) = 10-3 μm Two main types of cells in the living world: Prokaryotic cell, primitive cell type, most important representatives of which are the bacteria (capsule, plasma membrane, no cell organelles, only forrunner of the cell nucleus: „pro-karyon”). Size ≤ 1 μm. Eukaryotic cell,complex structure, real cell nucleus („eu-karyon”), cell organelles, membrane-bound cytoplasmic compartments, average size: 10-20 μm. The overwhelming majority of living organisms (including the human organism) consists of eukaryotic cells. Uni- and multicellular organisms. Cytology:a classical science dealing with cells, primarily with its structural aspects. Cell biology:an integrative science, which summarizes and integrates all our knowledge about structure, molecular composition, biochemistry, biophysics, physiology, pathological aspects of cells. Integration of structure and function. Teaching cell biology at our university.
Size difference between pro- and eukaryotic cells Nucleus of the eukaryotic cell prokaryotic cell (phagocyted bacterium) Mitochondrium (cell organelle of the eukaryotic cell) Macrophage (eukaryotic cell) with phagocyted bacteria (prokaryotic cells)
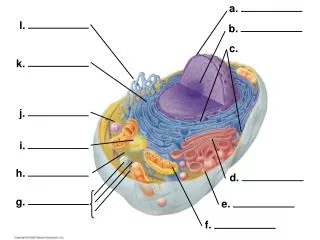
Components of an idealized animal cell (light and electron microscopic „inventory”) microvilli kinocilium intermediate filaments stereocilium phagocytosis cortical actin filaments phagosome nuclear envelope lysosome peroxisome mitochondrium secretory granule nucleus chromatin exocytosis microtubule nucleolus Golgi-apparatus lipid droplet pinocytosis coated pit glycogen granules centrosome smooth endoplasmic reticulum (sER) rough endoplasmic reticulum (rER)
Structural components of an animal cell Membrane-delimited compartments (ER, Golgi, lysosomes, mitochondria, transport vesicles, peroxisomes, …) plasma membrane cell nucleus Cell organelles protoplasm Not membrane-bound organelles (cytoskeleton, ribosomes) Reserve nutrients (inclusions) cytoplasm cytosol
The plasma membrane Microscopic morphology Each cell is covered by an extremely thin membrane that separates and protects the interior of the cell from its environment. At the same time, this so-called plasma membrane (or cell membrane) makes interaction of the cell with its environment possible and integrates the cell into its surroundings in multicellular organisms. In the light microscope the plasma membrane usually can not be seen due to its extreme thinness. Isolated erythrocyte membranes (after burst of the red blood cell and outflow of its contents in highly hypotonic solutions) can be just discerned as so-called „ghosts” in the phase-contrast microscope. In the electron microscope the plasma membrane can be seen as a fine black line, which can be resolved with high magnifications into a trilaminar structure (two dense layers with a light layer between them). Total thickness: 8-10 nm. plasma membrane High EM magnification Low EM magnification
Intracellular membranes Membranes are present also in the cell interior (intracellular membranes), where they form imortant constituents of a number of cell organelles (endoplasmic reticulum, Golgi-apparatus, lysosome, mitochondria, etc.). rER mito-chondrium Golgi-apparatus
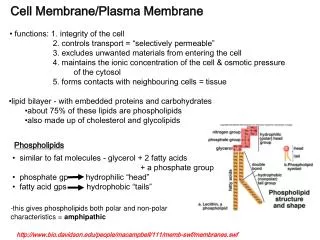
Basic structure of the membrane: the lipid bilayer I. Lipids Lipids are fat-like molecules with the largest part of the molecule being hydrophobic (water-repellent). However, these molecules also carry a hydrophilic (water-attractant) region or chemical group (amphiphilic molecules). Their main representatives in the membrane are: phospholipids, cholesterol and a few other lipid species. An example for phospholipids: phosphatidylcholin Cholesterol
Arrangement of lipids in an aqueous environment micelle bilayer Arrangement of phospholipids and cholesterol in the lipid bilayer Lipids occur in an aquous environment in two energetically stable supramolecular arrangements: the planar bilayer and the spherical micelle
Important physical properties of the lipid bilayer Diffusion barrier a barrier for hydrophilic molecules and ions across the membrane due to the continuous hydrophobic layer in the interior. However, it is … not a barrierfor hydrophobic molecules (e.g. O2, CO2, steroid hormones). The small water molecules can pass through the lipid bilayer with some difficulty. Dynamic structure: the bilayer behaves as a two-dimensional fluid. Mobility: lipid molecules are in continuous motion in the bilayer, they rotate around their long axis, their fatty acid chains are mobile, and Lateral diffusion: they diffuse in the plane of the bilayer with high velocity. However, a flip-flop translocation of lipids from one lipid layer into the other one is very rare and is only possible with the aid of special enzymes in the living cell. Fluidity of the bilayer structure depends on various factors , like relative amount of cholesterol (rigid molecule!), unsaturated fatty acids in the phospholipid molecules and temperature. Plasticity: Due to its dynamic structure, the lipid bilayer (the plasma membrane) is able to follow form and volume changes of the cell. Lipid asymmetry in the plasma membrane. Various types of phospholipids are asymmetrically distributed in the two layers of the membrane (e.g. phosphatidylcholin in the outer layer, phosphatidylserine and phosphatidylinositol in the inner layer).
Vesicles The lipid bilayer forms a spherical vesicle.Due to energetic reasons sheet-like (planar) bilayers do not occur in an aquous environment. Tears in the bilayer are quickly repaired by a spontaneous rearrangement of the lipids , vesicles are therefore quickly closed Fusion of lipid vesicles:vesicles can fuse with each other or with the plasma membrane (role in vesicular transport between membrane-limited cell organelles). Artificial lipid vesicles: liposomes(application in medicine. Import of drugs into cells). Liposomes visualized in a freeze-fracture sample. Electron micrograph
„Intelligent” properties of the membrane depend on membrane proteins II. Proteins Some basic knowledge about protein structure. Proteins are the most important molecules of life. Amino acids, polypeptide chain, 20 different side chains, 3D structure (folding), depending on the amino acid sequence, tertiary structure and function, conformational change and its significance. 25-75% of the membrane mass consists of proteins. About 50 lipid molecules per one protein. Association of proteins with the lipid bilayer Integral and peripheral membrane proteins 1-6 integral membrane proteins 7-8 peripheral membrane proteins
1. Integral membrane proteins form integral constituents of the membrane, are bound firmly to the lipid bilayer and can not be removed without destructing the bilayer Transmembrane proteinsextend across the bilayer, the polypeptide chain traversing the membrane once or many times („single-pass” and „multipass” proteins). The membrane-spanning portions are α-helixcal structures, where the hydrophobic side chains are turned outwards and are in hydrophobic interaction with the fatty acid chains of phospholipids. Most important membrane proteins (e.g. transport proteins, membrane receptors, cell adhesion proteins) belong to this group. Membrane proteins partially embedded into the bilayer These are similar to transmembrane proteins but do not span the bilayer. Their hydrophobic region is embedded into the cytosolic lipid monolayer (e.g. caveolin) Anchored membrane proteins are located on one side of the bilayer and are attached to it by a covalently bound lipid chain on the cytosolic side or by an oligosaccharide chain covalently bound to phosphatidyl inositol on the extracellular side (GPI anchor). The anchor can be detached from the protein (functionally important separation of the protein from the membrane) or, by binding back again to the anchor, reattached to the membrane. caveolin GPI-anchor fatty acid anchor
peripheral membrane protein 2. Peripheral membrane proteins These are hydrophilic proteins on either the extracellular or cytosolic surface of the membrane and are attached by weak bonds to an integral membrane protein. They can be removed from the membrane without destruction of the lipid bilayer. integral membrane protein III. Sugar constituents: Glycocalyx oligosaccharide chains peripheral glycoprotein At the extracellular surface of the cell membrane there are short sugar chains (oligosaccharide chains), covalently bound to proteins or lipids (glycoproteins, glycolipids). Some proteins carry long chains consisting of specific sugar molecules (glycosaminoglycan chains, GAGs), they are called proteoglycans. This carbohydrate-rich layer on the external (extracellular) side of the membrane is the glycocalyx. Its functional significance is only partially known (protection of the cell against bacteria, binding sites of membrane receptors, blood group antigens in erythrocytes, binding sites to other cells, etc.) The glycocalyx is rich in negatively charged (acidic) chemical groups and therefore can be visualized by applying positively charged colloidal stains (e.g. ruthenium red) as a dense layer in the electron microscope. GAG chains glycocalyx membrane
Membrane skeleton The membrane, consisting of the lipid bilayer and membrane proteins, is very thin and fragile and needs mechanical support. This consists of a network of filamentous proteins that is attached to the membrane by binding to integral membrane proteins. The network is called membrane skeleton. A further support for the membrane is the marginal, densely packed zone of the cytoskeleton (cell cortex, mainly consisting of actin filaments) that is also bound to the membrane, directly with adaptor proteins (like vinculin, talin, actinin) or indirectly. Membrane skeleton of red blood cells. A network of long spectrin and short actin filaments, bound to integral membrane proteins on the cytosolic surface of the cell membrane by specific proteins (ankyrin, band 4.1 protein). It maintains the elasticity and biconcave shape of the red blood cells. (its detachment from the membrane leads to loss of the biconcave shape and fragmentation of the erythrocytes into spherical structures). Cytosolic surface of the erythrocyte membrane with the membrane skeleton Membrane skeleton in other cell types: variations of the membrane skeleton in red blood cells, where spectrin is substituted with other, spectrin-related proteins, like fodrin.
Mobility of membrane proteins, membrane domains • Lateral diffusion: proteins are usually mobile and move in lateral direction in the plane of the lipid bilayer. • Exceptions: • 1. Membrane macrodomains • proteins can be immobilized by binding to stable structures outside the membrane (on one or the other side) or when they are bound to each other. • In the case of tight junction (see later) a linear arrangement of special membrane proteins in the lipid bilayer can form a barrier (fence) against lateral diffusion of proteins and can separate membrane regions with different protein and lipid compositions. axonal domain apical domain basolateral domain Examples(important in the specific function of certain cell types): apical and basolateral membrane domains in eithelia Dendritic and axonal membrane domains in nerve cells 2. Membrane microdomainsare small areas in the membrane with a molecular composition different from that of the surrounding membrane regions. lipid rafts: small membrane spots enriched in special lipids and proteins. Caveolae: lipid rafts with the integral membrane protein caveolin on the cytosolic side of the membrane. Vesicle-like invaginations of the membrane with various functions. dendritic domain epithelial cell nerve cell caveola
Functions of the cell membrane I. Diffusion barrier Basis of the barrier: the continuous hyrophobic layer in the interior of the lipid bilayer. It inhibits free diffusion of many substances across the membrane and makes thereby a controlled transport through the membrane possible. The membrane is a barrier against free diffusion of smaller or larger polar molecules as well as electrically charged particles (ions) It is not a barrier for hydrophobic molecules (e.g. gases, like O2, CO2, and steroid hormones). If the barrier is not functioning, e.g. by stripping, permeabilisation or poration of the membrane, the stable inner environment of the cell can not be maintained and the cell dies. Example: the natural killer cells of the immune system kill foreign cells by inserting pores into the membrane of the target cell.
II. Controlled uptake and release of substances through the membrane A completely closed system is not viable, the cell must have an interaction with its environment to survive. It has to take up nutrients and waste products have to be released. Comparison with a medieval town : city wall with controlled gates Two mechanisms for uptake and release: • Membrane transport:Transport of small molecules and ions across the membrane. 2. Endocytosis and exocytosis:Uptake and release of macromolecules or colloidal particles with vesicles (see later).
Transport across the membrane by transmembrane proteins. 20-30% of proteins in the cell are membrane transport proteins! Two large groups of transport proteins: transporter (carrier) proteins and channel proteins. kinn 1. Transporter (carrier) proteins A transporter protein transports a molecule or an ion by conformational change (binding of the molecule to be transported on one side, conformational change, release of the molecule on the other side of the membrane). lipid bilayer concentration gradient molecule ion Cotransport: 2 molecules or ions are simultaneously transported. A. Facilitated diffusion Molecules (or ions) are transported along their concentration gradient (from higher concentration towards lower concentration). This transport does not need energy (passive transport). COTRANSPORT
B. Active transport (pump) The transporter (carrier) protein transports a substance (e.g. ion) against its concentration gradient. This transport needs energy (active transport). Two energy sources: ATP-hydrolysis or potential energy of a concentration gradient. 1. ATP-driven pumps. Examples: Na+-K+-pump. Na+ are transported outwards and K+ inward, both against their concentration gradients, using the energy of ATP-hydrolysis. ABC transporter: pumps hydrophobic molecules from the cell into its environment with the hydrolysis of 2 ATP molecules. Example: in cancer chemotherapy the cancer cell can pump anti-cancer agents out from the cell. 2. Secondary active transport.Concentration gradient as energy source. Example: potential energy of water in a water tower. The energy for the transport comes from backflow of molecules or ions along their concentration gradient. Example: Na+ glucose transporter. electro-chemical gradient of Na glucose gradient
2. Channel proteins a) Ion channels • These proteins form a hydrophilic channel across the membrane, through which ions can flow from one side of the membrane to the other side. Ion selectivity(e.g. K-channels, Na-channels). Passivetransport. Rapid ion flow through the channel: thousend- times faster than with a transporter (in one second up to 1 million ions can flow through a channel). • Significance: primarily in uptake and conduction of electric impulses in nerve and muscle cells. • The open state of the channel is usually regulated. Various channels can be opened upon specific stimuli by conformational changes: • Ligand-gated channels: a signal molecule is bound to a specific binding site of a channel protein (e.g. neurotransmitters, nucleotides, ions). • Mechanically (stress)-gated channels react to mechanical stimuli (e.g. pressure on or stretch of the membrane). • Voltage-gated channels react to changes of the electric field in the immediate vicinity (important in nerve conduction). b) Water channels Aquaporin proteins with a water channel in their interior facilitate an intensive water flow through the membrane (e.g. kidney cells). Specific for water, no ions can flow through. c) Non-specific channels Porin-channelsin the outer mitochondrial membrane, Connexons in gap junction between two cells.
III. Contact between the cell and its environment The cell surface is a contact surface between the cell and its environment (important in the adaptation of the cell to its environment and in its integration in a multicellular organism). The cell surface is the „face” of the cell to the outside world. • 1. Signal registration.Membrane receptors(transmembrane proteins) that by binding and reacting to specific (chemical, physical, etc.) signals, switch on signal transduction pathways that lead informations into the interior of the cell. Receptor specificity. • 2. Cell-cell recognition.Adhesion molecules (transmembrane proteins) in the membrane are responsible for specific cell associations or for binding cells to the extracellular matrix. Important in the development of tissues and organs. • „Homing” of lymphocytes in lymphatic organs. Cell-cell contacts in the immune response. MHC complex (cell identity). • 3. Stable binding between cells:intercellular junctional structures (desmosome, tight junction, gap junction, … see later).
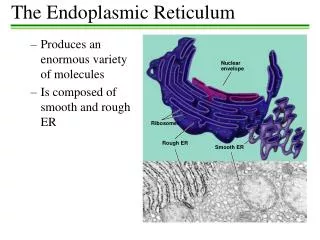
Endoplasmic reticulum (ER) Structure: tubular and/or flattened sac-like elements (ER tubules andcisternae), forming a 3D anastomosing network in the cytoplasm. These structural elements consist of a limiting membrane and of an internal space enclosed by the membrane. Discovered by Porter and Palade in the 1950s with the electron microscope. Two forms:rough ER: (ribosomes attached to the cytosolic surface of the membrane) and smooth ER. electron micrograph
Rough endoplasmic reticulum (rER) Less developed form Network composod of a few cisternae and tubules. Detail of a lutein cell. Highly developed form Many cisternae arranged parallelly to each other (also called ergastoplasm). Detail of a pancreatic acinar cell. EM EM
Function of the rER Main function: synthesis, translocation through the membrane and modification of certain proteins (secretory proteins, lysosomal enzymes, transmembrane proteins, resident proteins) Cotranslational transport (vectorial translation) • Translation begins on a cytosolic ribosome, the start sequence (15-35 amino acids) functions as a localisation signal (signal sequence). • The signal sequence is recognized and bound by a ribonucleoprotein complex (signal recognition particle: SRP) that blocks further translation. • SRP itself functions as a signal molecule and is bound to its receptor in the ER membrane (SRP-receptor, an integral membrane protein). thereby is the SRP-SRP receptor complex anchored to the ER membrane. • SRP is released and translation is continued. The forming peptide chain is sliding through a canal complex through the ER membrane, while the ribosome remains attached to the membrane • The signal sequence is cut off by an enzyme (signal peptidase) and the canal complex is disassembled into its subunits. signal sequence 1 canal 2 3 4
Termination of the cotranslational transport 1. Secretory and lysosomal proteinsare set free in the ER lumen 2. Integral membrane proteins remain partially embedded in the ER membrane, due to their hydrophobic stop-transfer signals (amino acid sequences) that anchor them in the membrane. A few protein modifications in the rER • Initial N-glykosylation (attachment of mannose-rich short sugar chains) • Hydroxylation (attachent of reactive OH grups onto certain proteins) • Formation of disulfide (S – S) bonds between parts of a polypeptide chain • Folding, quality control, correction with chaperones.
Smooth endoplasmic reticulum (sER) 3D interconnected system of tubules and cisternae. The large intracytoplasmic membrane area can accomodate various membrane-bound enzymes (and/or transport molecules). Detail of a retinal pigment epithelial cell. Fine tubules of the sER form a branching, labyrinthic system. Electron micrograph. Detail of a lutein cell. Dense network of cisternae of the sER. Electron micrograph.
Functions I. General function. Lipid synthesis. Enzymes for synthesis of phospholipids, cholesterol, neutral fat (triacylglycerol) are situated in the membrane system of the sER. Origin of the lipid bilayer. II. Special functions in certain cells.. • Glucose-6-phosphataseenzyme is located in the sER. This enzyme cleaves the phosphate group from glucose-6-phosphate (Glc-6-P) by which glucose can exit from the cell (example: liver cell). • Ca-storage.Ca-ATPase pumps Ca from the cytosol into the interior of the sER, where it is bound to Ca-binding proteins (e.g. calsequestrin). Upon specific signals Ca-ions are released through ligand-gated Ca-channels into the cytosol, where Ca-ions play important role in signal transduction pathways. A specialised sER is present in the striated muscle fiber: sarcoplasmic reticulum (important regulatory function in muscle contraction). • Retinal-reisomerisation.The retinal component of the photoreceptor molecule is reisomerised in the highly developed sER of the pigment epithelial cell from the trans isomeric form into the cis form (the cis form can absorb photons, regeneration of the photoreceptor molecule). The enzyme responsible for reisomerisation (isomerohydrolase) is localised in the sER membrane. • Synthesis and modification of steroid hormones (p450 monooxygenases). In: cells of the adrenal cortex, Leydig-cells of the testis, lutein cells of the ovary. • Detoxication.Hydrophobic molecules (e.g.. xenobiotics, that act as toxins) accumulating in fat and lipids of the organism are made hydrophilic by addition of polar groups (OH groups, glucuronic acid, etc.) by the enzymes p450 monooxigenases. The toxins this way can be excreted and eliminated from the organism. A typical detoxicating cell: the liver cell.
Textbook: Alberts: Essential cell biology, Garland Science ed., New York, London Chapter 1: Eukaryoticcell Chapter 11: Membranestructure (Lipidbilayer, membraneproteins) Chapter 12: Membranetransport (Transporters, Ion channels) Chapter 15: Intracellularcompartments (Membrane-enclosedorganelles, protein sorting) + lecture, (summarized in the present ppt presentation). Sources of figures Röhlich: Histology, 4th. edition, Semmelweis Publishing House, Budapest, 2014 (in Hungarian) Alberts – Johnson – Lewis – Raff – Roberts – Walter: Molecular biology of the cell. 5th edition, Garland Science Specimens, micrographs and drawings of P. Röhlich Campbell – Reece: Biologie, Spektrum - Fischer