Download
1 / 22
360 likes | 1.12k Views
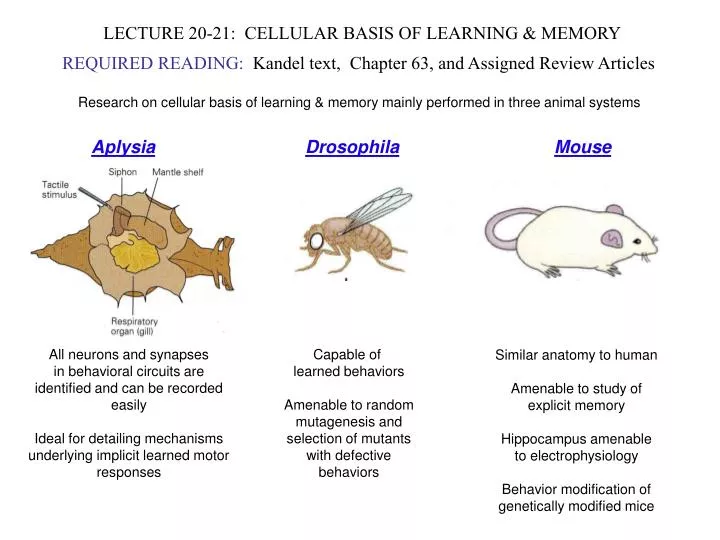
LECTURE 20-21: CELLULAR BASIS OF LEARNING & MEMORY. REQUIRED READING: Kandel text, Chapter 63, and Assigned Review Articles. Research on cellular basis of learning & memory mainly performed in three animal systems. Aplysia Drosophila Mouse. All neurons and synapses

E N D
LECTURE 20-21: CELLULAR BASIS OF LEARNING & MEMORY REQUIRED READING: Kandel text, Chapter 63, and Assigned Review Articles Research on cellular basis of learning & memory mainly performed in three animal systems AplysiaDrosophilaMouse All neurons and synapses in behavioral circuits are identified and can be recorded easily Ideal for detailing mechanisms underlying implicit learned motor responses Capable of learned behaviors Amenable to random mutagenesis and selection of mutants with defective behaviors Similar anatomy to human Amenable to study of explicit memory Hippocampus amenable to electrophysiology Behavior modification of genetically modified mice
APLYSIA SHORT-TERM LEARNED RESPONSES AFFECTING GILL WITHDRAWL REFLEX CLASSICAL CONDITIONING HABITUATION SENSITIZATION Harmful stimulus sensitizes gill withdrawl response to subsequent harmful OR harmless stimuli given to same OR different body regions Pairing harmful stimulus with preceding harmless conditioning stimulus sensitizes gill withdrawl response to subsequent conditioning stimulus but not to tactile stimuli given to other body areas Repeated tactile stimulation of siphon depresses gill withdrawl response
HABITUATION IS DUE TO DEPRESSED NEUROTRANSMITTER RELEASE AT SEVERAL SITES Rapidly repeated tactile stimulation of siphon attenuates gill withdrawl both during the training and for a short period afterwards. Habituation is due to reduced neurotransmitter release by the sensory neuron and by relevant interneurons in response to the tactile stimulus. I.e., the memory of habituation is distributed at various synapses in the circuit Whereas a rapid series of stimuli induces short-term habituation, several sets of tactile stimuli distributed over several hours induces long-term habituation that lasts for weeks. Long-term habituation requires new protein synthesis and is due to pruning of synaptic connections
SHORT-TERM SENSITIZATION IS MEDIATED THROUGH AXO-AXONIC SEROTONERGIC SYNAPSES OF FACILITATING INTERNEURONS Serotonergic facilitating interneurons send axo-axonic connections to broadly distributed sensory neurons Unconditioned stimulus causes interneurons to release serotonin, which acts through metabotropic HT receptors to increase the likelihood of neurotransmitter release following sensory neuron firing Sensitization can be mimicked without sensitizing stimulus by local experimental application of serotonin Sensitization is mediated by presynaptic elevation of cAMP & PKA activity, which has three effects: Greater proportion of vesicles in active zone (synapsin phosphorylation?) K+ channel inactivation increases duration of depolarization and magnitude of Ca+2 influx 3) Activation of L--type calcium channels
CLASSICAL CONDITIONING EMPLOYS SEQUENCE-REINFORCED PRODUCTION OF cAMP Conditioning is only effective when CS precedes US by a short interval (~ 0.5 sec) CS elevates calcium in presynaptic terminal at moment of US. Calcium/CAM enhances the enzymatic activity of adenylate cyclase triggered by 5-HT. Adenylate cyclase is a biochemical “coincidence detector”
TEMPORALLY SPACED SENSITIZATION OR CONDITIONING TRAININGSINDUCE LONG-TERM IMPLICIT MEMORY Long-term sensitization and conditioning are also mediated through presynaptic cAMP production and PKA activity PKA induces specific CREB-dependent gene transcription and protein synthesis: Newly synthesized ubiquitin hydrolase degrades PKA regulatory subunits, making the enzyme constitutively active Other newly synthesized proteins help build new presynaptic terminals onto motor neurons
GENETIC SCREENS FOR GENES AFFECTING CONDITIONING IMPLICIT MEMORY ALL AFFECT THE cAMP-PKA-CREB PATHWAY FLY MUTANTS SELECTED FOR DEFECTS IN IMPLICIT MEMORY DUNCEencodes cAMP phosphodiesterase RUTABAGA mutant defective for Ca+2/CAM enhancement of cyclase AMNESIACencodes a peptide neurotransmitter acting on GS-coupled receptor PKA-R1encodes PKA
HIPPOCAMPAL NEURONS IN DIFFERENT RELAYS ARE ALL CAPABLE OF UNDERGOING SYNAPTIC LONG-TERM POTENTIATION One Theta burst gives what is sometimes called Early LTP, which is less than doubling of EPSC which lasts for hours Four Theta bursts spaced minutes apart generate Late LTP, with up to 4-fold EPSC stimulation that lasts for days AMPLITUDE OF EPSCS AXON STIMULATION PROTOCOL 300 Once per minute Once per minute 10 Hz 200 EPSP Slope (% original) 20 min 1 m 60 min 100 20 40 60 80 “THETA” BURST TIME (min)
INDUCTION AND EXPRESSION OF SYNAPTIC PLASTICITY Prior synaptic activity can INDUCE long-term plasticity. Such plasticity can be INDUCED by molecular events occuring either presynaptically or postsynaptically. The changes in transmission following synaptic plasticity can be EXPRESSED either presynaptically and/or postsynaptically, and need not correspond to the site of INDUCTION. E.g., at a certain synapse, postsynaptic calcium influx can INDUCE plasticity which is then EXPRESSED as changes in presynaptic neurotransmitter release probability.
LTP AT MOSSY FIBER--CA3 SYNAPSES IS DUE TO PRESYNAPTIC CALCIUM INFLUX AND cAMP/PKA PATHWAY
LTP AT SCHAFFER COLLATERAL--CA1 SYNAPSES IS DUE TO POSTSYNAPTIC CALCIUM INFLUX AND CAM KINASE ACTIVITY LTP at CA3-CA1 synapse is blocked by NMDAR antagonist APV and by inhibitors of CAM kinase
PRESYNAPTIC COMPONENT OF EARLY AND LATE LTP AT CA3--CA1 SYNAPSES RESEMBLES SHORT- AND LONG-TERM SENSITIZATION Late LTP absolutely requires new protein synthesis
PRESYNAPTIC COMPONENT OF EARLY AND LATE LTP REQUIRES POSTSYNAPTIC CAMK ACTIVITY AND RETROGRADE SIGNALS
OTHER MECHANISMS OF PLASTICITY ENHANCING EPSPS LTP can be expressed postsynaptically as a reduction of leak conductance in dendritic spine. This enables the EPSC to generate EPSP with greater length and time constants. Excitatory transmission can be enhanced by HETEROSYNAPTIC INHIBITION OF INHIBITORY TRANSMISSION. This is mediated by endogenous cannabinoids acting on presynpatic terminals of nearby GABAergic synapses.
IS LTP REQUIRED FOR HIPPOCAMPAL CONSOLIDATION OF EXPLICIT MEMORY? CAMK AND NMDAR1 NEEDED FOR LONG-TERM SPATIAL REPRESENTATION IN HIPPOCAMPUS Single pyramidal neuron in hippocampus fires when mouse is in certain location (independent of animal’s orientation) Normal mouse remembers where it has been. spatial map in HC does not change in subsequent chamber trials Mice with hippocampus- restricted mutations in CAMK or NMDAR1 establish place fields, but do not remember from day to day
IS LTP REQUIRED FOR HIPPOCAMPAL CONSOLIDATION OF EXPLICIT MEMORY? HIPPOCAMPAL CAMK AND NMDAR1 NEEDED FOR BOTH LTP AND SPATIAL MEMORY
SYNAPSES SENSITIVE TO NMDAR-MEDIATED LTP ARE ALSO SENSITIVE TO NMDAR-MEDIATED LONG-TERM DEPRESSION (LTD) AMPLITUDE OF EPSCS AXON STIMULATION PROTOCOL 300 LTP Once per minute Once per minute 10 Hz 200 EPSP Slope (% original) 20 min 1 m 60 min 100 20 40 60 80 TIME (min) 300 200 EPSP Slope (% original) Once per minute Once per minute LTD 100 2 Hz 20 min 5 m 60 min 20 40 60 80 TIME (min)
LTD HAS A LOWER CALCIUM CONCENTRATION THRESHOLD THAN LTP, BUT LTP IS DOMINANT THETA- OR HIGH-FREQUENCY STIMULUS TRAIN GREATER CALCIUM ENTRY ACTIVATION OF CALCINEURIN AND CAMK AMPA RECEPTOR INSERTION AND PHOSPHORYLATION LTP LOW-FREQUENCY STIMULUS TRAIN LOW-LEVEL CALCIUM ENTRY ACTIVATION OF CALCINEURIN AMPA RECEPTOR INTERNALIZATION LTD
STRUCTURAL AND FUNCTIONAL FEATURES OF AMPA-TYPE GLUTAMATE RECEPTORS AMPA receptors are homo- or hetero-tetramers Restriction of calcium entry mediated by GluR2; tetramers containing >1 GluR2 subunit conduct only Na+/K+ AMPA receptors encoded by different genes or by alternative splicing have different C-terminal tails. Receptor tails contain phosphorylation sites for different protein kinases and binding sites for PDZ-domain-containing proteins Receptors containing only GluR2(short) and/or GluR3 subunits are delivered constitutively from vesicles to synapse Retention at synapse mediated by complex with Glutamate Receptor Interacting Protein (GRIP) Receptors containing at least one GluR1(long) subunit are stored in intracellular vesicles near synapse During LTP, GluR1-containing tetramers are added to the synapse
NMDAR-INDUCED CAMK ACTIVITY ACTS ON AMPA RECEPTORS IN TWO WAYS TO PROMOTE LTP CAMK phosphorylates an unknown protein, enabling a PDZ-protein that interacts with long tail on GluR1 to deliver receptor TO EXTRASYNAPTIC SITE Delivered receptors migrate (randomly?) into post-synaptic density, where interactions of receptor- associated GRIP and STG and the major postsynaptic matrix protein PSD-95 anchor receptor to synapse Newly delivered GluR1-containing AMPA receptors can be phosphorylated directly by CAMK, which increases unitary conductance of the receptor STG GRIP PSD-95 GRIP PSD-95 CAMK PDZ-protein Calcineurin Calcineurin activation promotes internalization of AMPA receptors containing only short-tail subunits, thereby promoting LTD WHEN HIGH CALCIUM ENTRY ACTIVATES BOTH CALCINEURIN AND CAMK, CAMK-MEDIATED GluR1-CONTAINING AMPAR EXOCYTOSIS EXCEEDS CALCINEURIN-MEDIATED SHORT TAIL-ONLY AMPAR ENDOCYTOSIS
HIGH CAMK ACTIVITY INDUCED DURING LATE LTP IS ALSO MEDIATED BY NEW CAM KINASE PROTEIN SYNTHESIS NEAR THE SYNAPSE Most mRNAs have 3’ polyA tail, which is necessary for initiation of the mRNA’s translation Neurons contain some mRNAs that are not polyadenylated, are not translated, and are transported along dendrites to areas near dendritic spines NMDA receptor activation and calcium entry activates a protein kinase called AURORA Aurora kinase activates translation of nearby dormant mRNAs ONE OF THESE DORMANT RNAs ENCODES CAM KINASE Because of its dendritic localizaation, new CAMK synthesis is restricted to the synapse undergoing LTP The dendritic localization of dormant CAMK RNA and its activation during LTP are mediated by Cytoplasmic Polyadenylation Element Binding (CPEB) protein
HOW DOES CPEB PROTEIN CONTROL RNA DORMANCY AND ACTIVATION IN NEURONS? PolyA is needed for assembly of 5’ translation initiation complex CPEB protein binding to 3’ CPE helps mask RNA 5’ end CPEB phosphorylation by Aurora allows for recruitment of polyA polyermerase (PAP) Polyadenylation of dormant RNA allows assembly of 5’ translation initiation complex