Download

1 / 55
600 likes | 932 Views
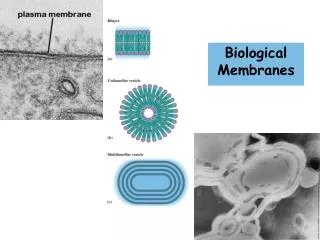
Chapter 12 Biological Membranes & Transport. Fluid Mosaic Model for Membrane Structure. Amphipathic lipid aggregates that form in water. or Vesicle. Bilayers are noncovalent, cooperative structures. Monolayer of oil molecules at an air-water interface. Membrane Phase Transitions.

E N D
Chapter 12 Biological Membranes & Transport
Amphipathic lipid aggregates that form in water or Vesicle Bilayers are noncovalent, cooperative structures
Membrane Phase Transitions The "melting" of membrane lipids • Below a certain transition temperature, membrane lipids are rigid and tightly packed • Above the transition temperature, lipids are more flexible and mobile • The transition temperature is characteristic of the lipids in the membrane
Higher the proportion of saturated fatty acid, higher is the transition temperature.
Sterol content of a membrane has 2 effects on membrane fluidity Below the transition temperature: Insertion of rigid planar sterol prevents highly ordered packing of fatty acid side chains Membrane fluidity Above the transition temperature: Rigid planar sterol reduces the freedom of neighboring fatty acid side chains Membrane fluidity
Cells regulate their lipid composition to achieve a constant membrane fluidity under various growth conditions
Motion of Membrane Lipids Lateral Diffusion Transbilayer or flip-flop Diffusion
Flippases A relatively new discovery! • Lipids can be moved from one monolayer to the other by flippase proteins • Some flippases operate passively and do not require an energy source • Other flippases appear to operate actively and require the energy of hydrolysis of ATP
Demonstration of lateral diffusion of membrane proteins Membrane proteins, like membrane lipids, are free to diffuse laterally in the plane of the bilayer
Restricted motion of the erythrocyte chloride-bicarbonate exchanger
Asymmetric distribution of phospholipids between the inner & outer monolayers of erythrocyte plasma membrane
Structure of Membrane Proteins Singer & Nicolson defined two classes • Integral (intrinsic) proteins • Peripheral (extrinsic) proteins • We'll note a new one – lipid-anchored proteins
Some membrane proteins span the lipid bilayer Glycophorin in the erythrocyte A single-transmembrane-segment protein • One transmembrane segment with globular domains on either end • Transmembrane segment is alpha helical and consists of 19 hydrophobic amino acids • Extracellular portion contains oligosaccharides (and these constitute the ABO and MN blood group determinants)
Lipid-linked membrane proteins Covalently attached lipids anchor membrane proteins to the lipid bilayer A relative new class of membrane proteins 4 types have been found: Amide-linked myristoyl anchors Thioester-linked fatty acyl anchors Thioether-linked prenyl anchors Glycosyl phosphatidylinositol anchors Glycosyl phosphatidylinositol (GPI) anchor
Integral Membrane Proteins Held in the membrane by hydrophobic interactions with lipids
3-D structure of the photosynthetic reaction center of purple bacterium First integral membrane protein to have its structure determined by X-ray diffraction methods Prosthetic group (light-absorbing pigments) Residues that are part of the trans-membrane helices
Porin FhuA, an integral membrane protein with b-barrel structure Not all integral membrane proteins are composed of transmembrane a helices Porin allows certain polar solutes to cross the outer membrane of bacteria
Porins Found both in Gram-negative bacteria and in mitochondrial outer membrane • Porins are pore-forming proteins (30-50 kD) • Most arrange in membrane as trimers • High homology between various porins • Porin from Rhodobacter capsulatus has 16-stranded beta barrel that traverses the membrane to form the pore
Why Beta Sheets? for membrane proteins?? • Genetic economy • Alpha helix requires 21-25 residues per transmembrane strand • Beta-strand requires only 9-11 residues per transmembrane strand • Thus, with beta strands , a given amount of genetic material can make a larger number of trans-membrane segments
Integral membrane proteins mediate cell-cell interactions & adhesion 4 examples of integral protein types that function in cell-cell interaction Serve as receptors & signal transducers Essential part of the blood-clotting process
Gap Junctions Vital connections for animal cells • Provide metabolic connections • Provide a means of chemical transfer • Provide a means of communication • Permit large number of cells to act in synchrony (for example, synchronized contraction of heart muscle is brought about by flow of ions through gap junctions)
Induces closure of gap junction central channel Gap Junctions • Hexameric arrays of a single 32 kD protein • Subunits are tilted with respect to central axis • Pore in center can be opened or closed by the tilting of the subunits, as response to stress
Cont. Chapter 12 Biological Membranes & Transport For chapter 12 Focus on the material covered in lectures Will not be tested on materials covered in Pages 424 - 429
Membrane fusion is central to many biological processes Membranes undergo fusion without losing its integrity
Movements of solutes across a permeable membrane Electrically neutral solutes Electric gradient or membrane potential
Energy changes accompanying passage of a hydrophilic solute through the lipid bilayer of a biological membrane Energy of activation Facilitated diffusion or passive transport
Aquaporins form hydrophilic transmembrane channels for the passage of water Proposed structure of aquaporin channel (Formed by 4 monomers) Likely transmembrane topology of an aquaporin, AQP-1 Monomer Water flows through the channel in single file at the rate of 5 X 108 molecules / second
Glucose transporter of erythrocytes mediates passive transport Monomer 1 12 Proposed structure of GluT1
A helical wheel diagram Shows the distribution of polar& non-polar residues on the surface of a helical segment
Side-by-side association of 5 or 6 amphipathic helices Polar
Model of glucose transport into erythrocytes by GluT1 T1 & T2 are 2 different conformations T1 has glucose binding site on the outer surface of the membrane T2, with the binding site on the inner surface
Three general classes of transport system • Differ in # of solutes • transported • & • 2) the direction in which each is transported
Types of transport Passive: Transported species always moves down its electrochemical gradient and it is not accumulated above the equilibrium point ATP not required Active:Results in accumulation of solute above the equilibrium point ATP is required
Na+K+ ATPase • In animal cells, this active transport system is responsible for • maintaining intracellular • Na+ and K+ concentrations • for generating transmembrane • electrical potential
Postulated mechanism of Na+ andK+ transport by the Na+K+ ATPase
Na, K Transport • Hypertension involves apparent inhibition of sodium pump. (Inhibition in cells lining blood) • Studies show this inhibitor to be ouabain!
A defective ion channel causes cystic fibrosis CF is the result of one amino acid change in the protein CFTR, a chloride ion channel Topology of cystic fibrosis transmembrane conductance regulator, CFTR